мотонейрон
- мотонейрон
-
- мотонейрон
-
сущ.
, кол-во синонимов: 1
Словарь синонимов ASIS.
.
2013.
.
Синонимы:
Смотреть что такое «мотонейрон» в других словарях:
-
мотонейрон — мотонейрон … Орфографический словарь-справочник
-
Мотонейрон — (лат. motor приводящий в движение + нейрон; син. нейрон двигательный) крупная нервная клетка в передних рогах спинного мозга. Мотонейроны обеспечивают моторную координацию и поддержание мышечного тонуса … Википедия
-
мотонейрон — (лат. motus движение, motor приводящий в движение + нейрон; син. нейрон двигательный) эфферентный нейрон, иннервирующий мышечные волокна … Большой медицинский словарь
-
мотонейрон — а, ч., анат. Будь яка окрема нервова клітина, яка активує ефектор … Український тлумачний словник
-
Мотонейрон — (motoneuron, от лат. motus движение, motor приводящий в движение + neuron нерв) – эфферентный нейрон, иннервирующий мышечные волокна … Словарь терминов по физиологии сельскохозяйственных животных
-
альфа-мотонейрон — мотонейрон передних рогов спинного мозга, аксон которого иннервирует экстрафузальные мышечные волокна … Большой медицинский словарь
-
Мотонейрон периферический — Непосредственно иннервирует мышечные волокна. Располагается в двигательных ядрах черепных нервов или в передних рогах спинного мозга … Энциклопедический словарь по психологии и педагогике
-
Мотонейрон центральный — Расположен в коре больших полушарий или в экстрапирамидных структурах головного мозга. Участвует в опосредованной иннервации мышц … Энциклопедический словарь по психологии и педагогике
-
гамма-мотонейрон — нейрон передних рогов спинного мозга, иннервирующий интрафузальные мышечные волокна … Большой медицинский словарь
-
Нейрон Двигательный, Мотонейрон (Motor Neurone) — нейрон (эффекторный), иннервирующий мышцу. Одни мотонейроны целиком расположены в центральной нервной системе. Их тела залегают в головном мозге (двигательной области коры полушарий большого мозга), а аксоны направляются к двигательным ядрам… … Медицинские термины
Нервная ткань — основная ткань, формирующая нервную систему и создающая условия для реализации ее многочисленных функций. Нервная ткань имеет эктодермальное происхождение, не принято делить нервную ткань на какие-либо виды тканей. Обладает двумя основными свойствами: возбудимостью и проводимостью.
Нейрон
Структурно-функциональной единицей нервной ткани является нейрон (от др.-греч. νεῦρον — волокно, нерв) — клетка с одним
длинным отростком — аксоном (греч. axis — ось), и одним/несколькими короткими — дендритами (греч. dendros — дерево).
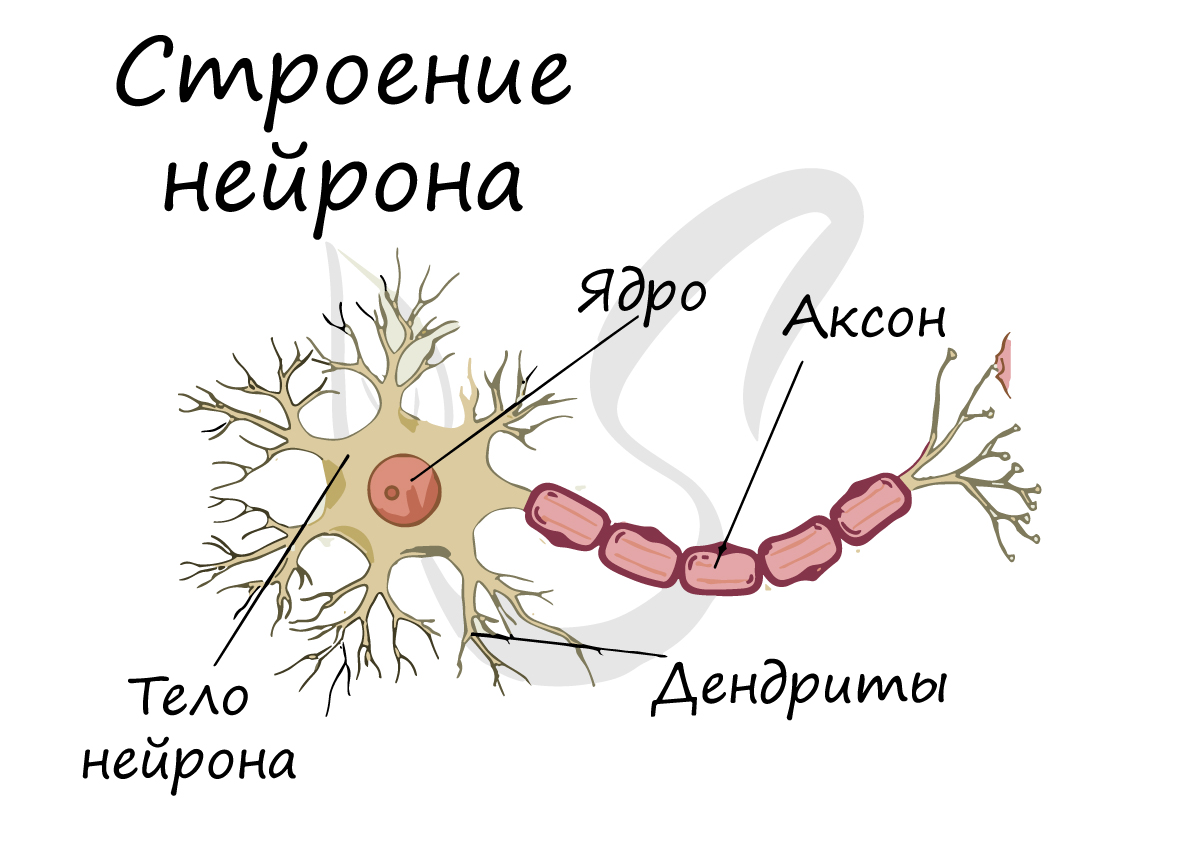
Спешу сообщить, что представление, будто короткий отросток нейрона — всегда дендрит, а длинный — всегда аксон, в корне неверно. С точки
зрения физиологии правильнее дать следующие определения: дендрит — отросток нейрона, по которому нервный импульс перемещается к телу нейрона, аксон — отросток нейрона, по которому импульс перемещается от тела нейрона.
Нейроны обладают 4 свойствами:
- Рецепция (лат. receptio — принятие) — способны воспринимать поступающие сигналы (дендриты)
- В ответ на сигналы способны переходить в состояние возбуждения или торможения
- Проведение возбуждения (от дендрита к телу нейрона, затем — к концу аксона)
- Передача сигнала другим объектам — нейрону или эффекторному органу
В физиологии эффекторным (от лат. efferes — выносящий) органом часто называют исполнительный орган или орган-мишень воздействия (мышцы, железы). Орган-эффектор выполняет те или иные «приказы» ЦНС (центральной нервной системы) или эндокринных желёз
Отростки нейронов проводят нервные импульсы и передают их другим нейронам, эффекторам, благодаря чему
мышцы сокращаются или расслабляются, а секреция желез усиливается или уменьшается.
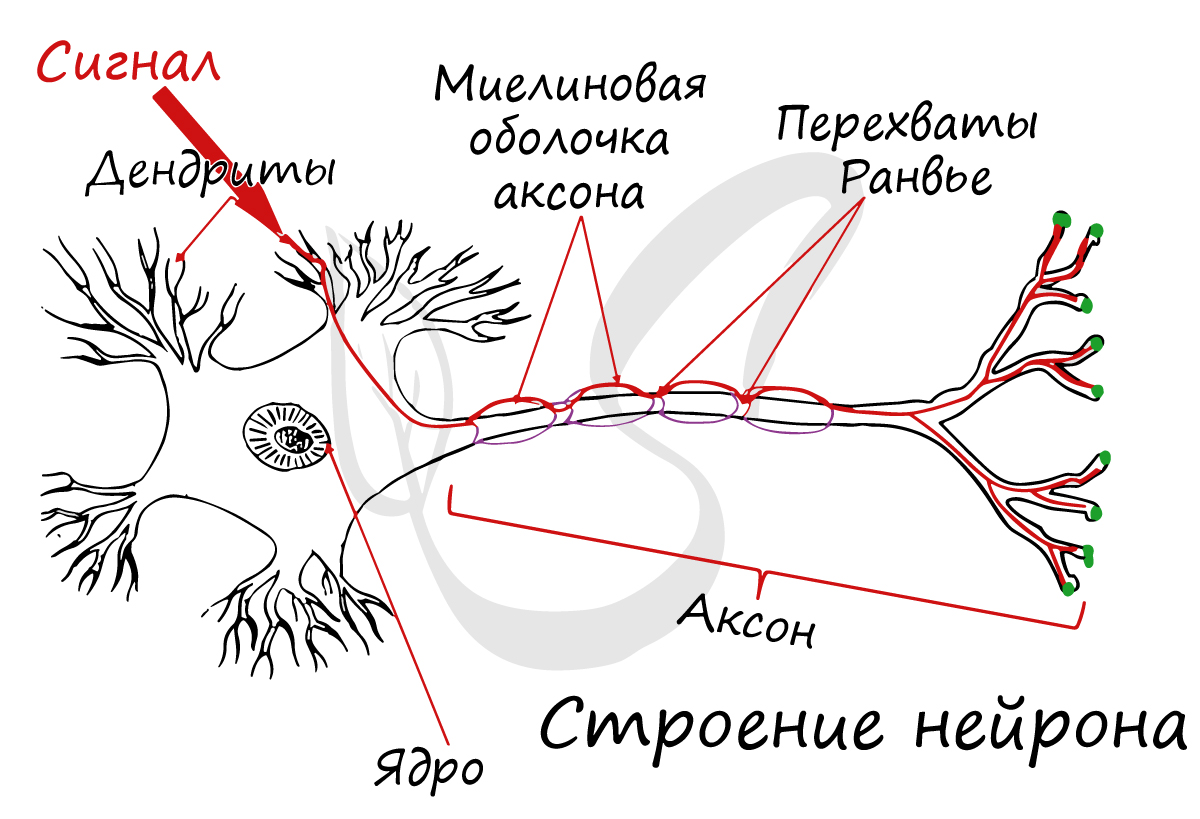
Миелиновая оболочка
Нервные волокна подразделяются на миелиновые и безмиелиновые. Нервное волокно — это один или несколько отростков нейронов (могут быть как аксоны, так и дендриты) с окружающей оболочкой.
Безмиелиновые нервные волокна находятся преимущественно в составе вегетативной нервной системы (скорость проведения 1-2 м/c). Миелиновые — образуют белое вещество головного и спинного мозга, нервные волокна соматической нервной системы (5-120 м/с).
В миелиновых нервных волокнах отростки нейронов покрыты миелиновой оболочкой (на 70-75% состоит из липидов (жиров)), которая обеспечивает изолированное проведение нервного
импульса по нерву. Если бы не было миелиновой оболочки (вообразите!) нервные импульсы распространялись бы хаотично, и,
когда мы хотели сделать движение рукой, то вместе с рукой двигалась бы нога.
Существует болезнь при которой собственные антитела уничтожают миелиновую оболочку нервных волокон головного и спинного мозга (случаются и такие сбои в работе организма). Эта
болезнь — рассеянный склероз, по мере прогрессирования приводит к разрушению не только миелиновой оболочки, но и нервов — а значит,
происходит атрофия мышц и человек постепенно становится обездвиженным.

Миелиновый слой представлен несколькими слоями мембраны глиальной клетки (леммоцит, шванновская клетка), которые закручиваются вокруг осевого цилиндра (отростка нейрона). Это закручивание хорошо видно на картинке, где изображен здоровый нерв, чуть выше 
Миелиновый слой оболочки волокна регулярно прерывается в местах стыка соседних леммоцитов — перехваты Ранвье. Миелиновая оболочка обеспечивает изолированное и более быстрое проведение возбуждения (сальтаторный тип, лат. salto — скачу, прыгаю).

Нейроглия (греч. νεῦρον — волокно, нерв + γλία — клей)
Вы уже убедились, насколько значимы нейроны, их высокая специализация приводит к возникновению особого окружения — нейроглии.
Нейроглия (глиальные клетки, глиоциты) — вспомогательная часть нервной системы, которая выполняет ряд важных функций:
- Опорная — поддерживает нейроны в определенном положении
- Регенераторная (лат. regeneratio — возрождение) — в случае повреждения нервных структур нейроглия способствует регенерации
- Трофическая (греч. trophe — питание) — с помощью нейроглии осуществляется питание нейронов: напрямую с кровью нейроны не контактируют
- Электроизоляционная — леммоциты (шванновские клетки) закручиваются вокруг отростков нейронов и формируют миелиновую оболочку
- Барьерная и защитная — изолируют нейроны от тканей внутренней среды организма
- Некоторые глиоциты секретируют цереброспинальную (спинномозговую) жидкость — ликвор (от лат. liquor — жидкость)
В состав нейроглии входят разные клетки, их в десятки раз больше чем самих нейронов. В периферическом отделе нервной
системы миелиновая оболочка, изученная нами, образуется именно из нейроглии — шванновских клеток (леммоцитов). Между ними хорошо
заметны перехваты Ранвье — участки, лишенные миелиновой оболочки, между двумя смежными шванновскими клетками.
Классификация нейронов
Нейроны функционально подразделяются на чувствительные, двигательные и вставочные.
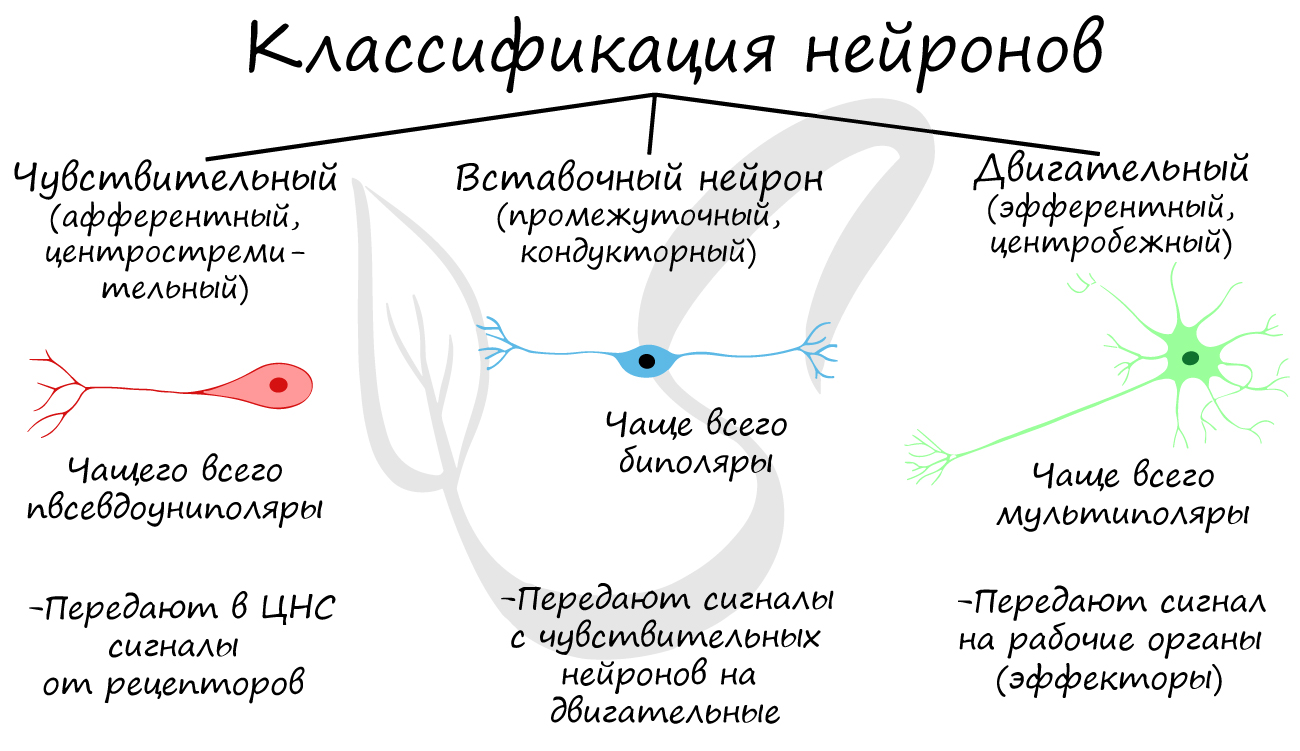
Чувствительные нейроны также называются афферентные, центростремительные, сенсорные, воспринимающие — они воспринимают раздражения, преобразуют их в нервные импульсы и передают в ЦНС. Рецептором называют концевое окончание чувствительных нервных
волокон, воспринимающих раздражитель.
Вставочные нейроны также называются промежуточные, ассоциативные — они обеспечивают связь между чувствительными и двигательными
нейронами, передают возбуждение в различные отделы ЦНС, участвуют в обработке информации и выработке команд.
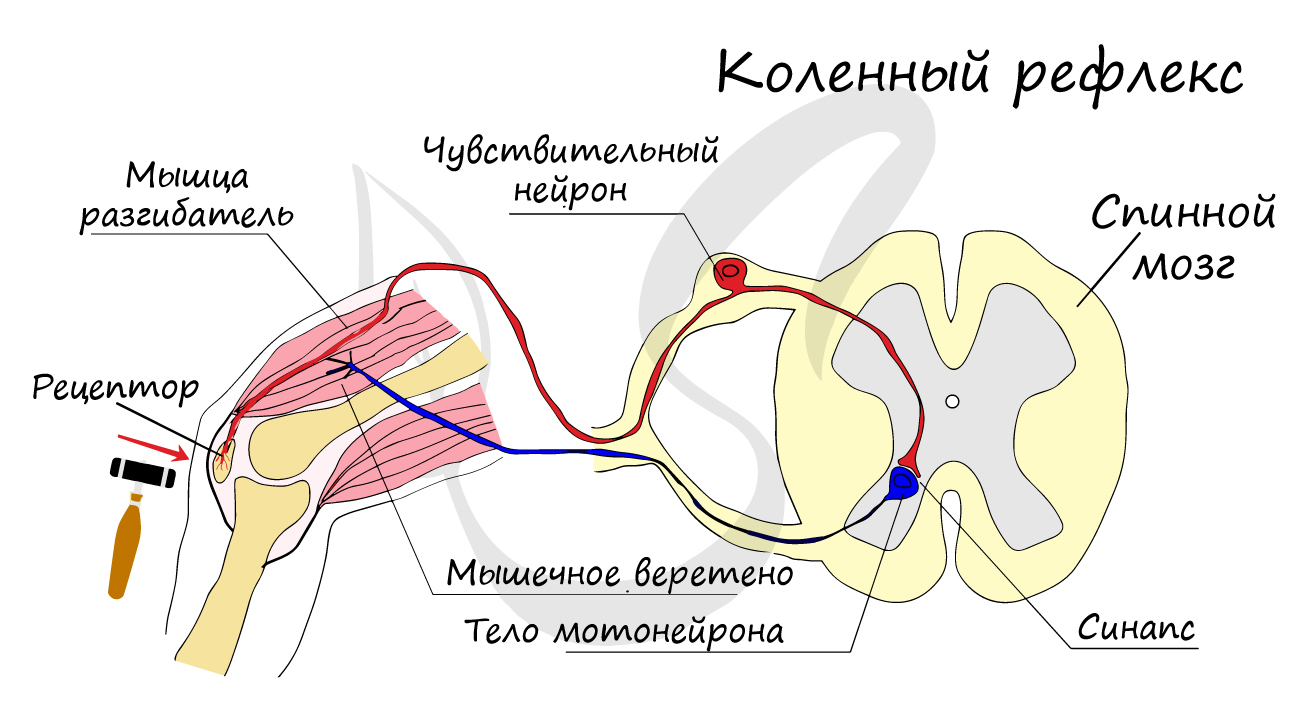
Двигательные нейроны по-другому называются эфферентные, центробежные, мотонейроны — они передают нервный импульс (возбуждение) на
эффектор (рабочий орган). Наиболее простой пример взаимодействия нейронов — коленный рефлекс (однако вставочного нейрона
на данной схеме нет). Более подробно рефлекторные дуги и их виды мы изучим в разделе, посвященном нервной системе.
Синапс
На схеме выше вы наверняка заметили новый термин — синапс (греч. sýnapsis — соединение). Синапсом называют место контакта между двумя нейронами или между
нейроном и эффектором (органом-мишенью). В синапсе нервный импульс «преобразуется» в химический: происходит выброс особых
веществ — нейромедиаторов (наиболее известный — ацетилхолин) в синаптическую щель.
Разберем строение синапса на схеме. Его составляют пресинаптическая мембрана аксона, рядом с которой расположены везикулы (лат. vesicula — пузырек) с
нейромедиатором внутри (ацетилхолином). Если нервный импульс достигает терминали (окончания) аксона, то везикулы начинают
сливаться с пресинаптической мембраной: ацетилхолин поступает наружу, в синаптическую щель.
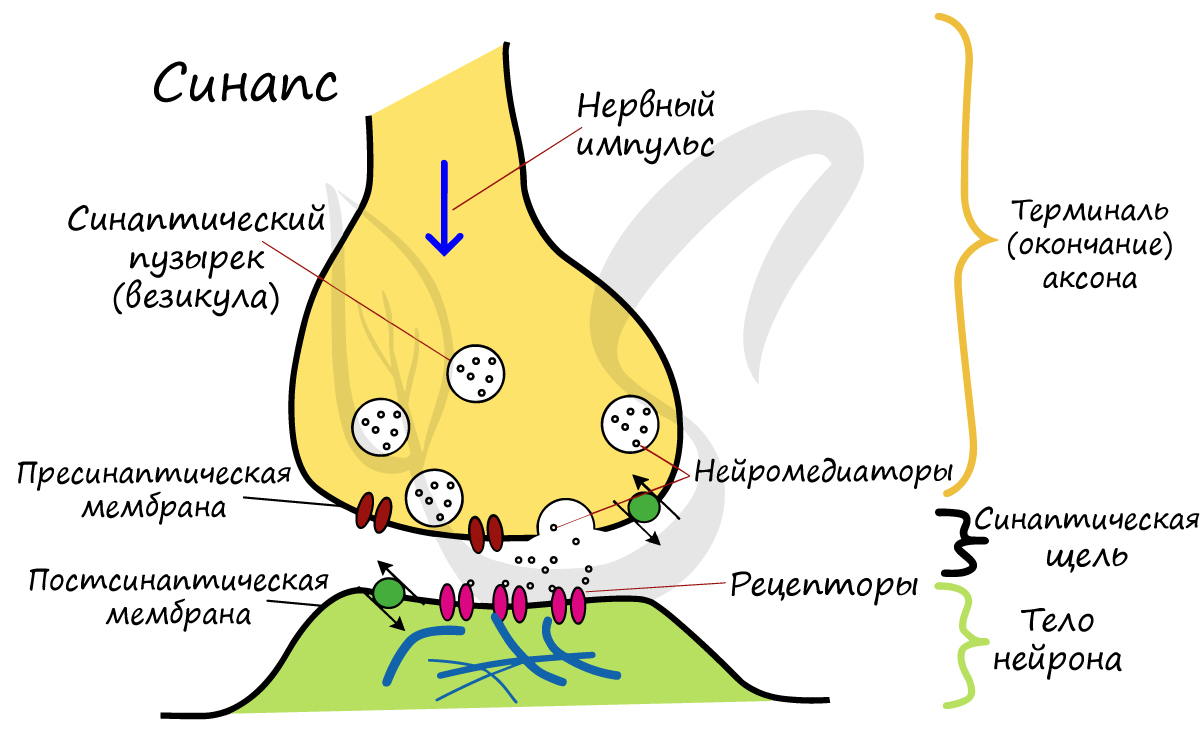
Попав в синаптическую щель, ацетилхолин связывается с рецепторами на постсинаптической мембране, таким образом, возбуждение (нервный импульс)
передается другому нейрону. Так устроена нервная система: электрический путь передачи сменяется
химическим (в синапсе).
Яд кураре
Гораздо интереснее изучать любой предмет на примерах, поэтому я постараюсь как можно чаще радовать вас ими Не могу утаить
историю о яде кураре, который используют индейцы для охоты с древних времен.
Этот яд блокирует ацетилхолиновые рецепторы на постсинаптической мембране, и, как следствие, химическая передача возбуждения с
одного нейрона на другой становится невозможна. Это приводит к тому, что нервные импульсы перестают поступать к эффекторам,
в том числе к дыхательным мышцам (межреберным, диафрагме), вследствие чего дыхание останавливается и наступает смерть животного.

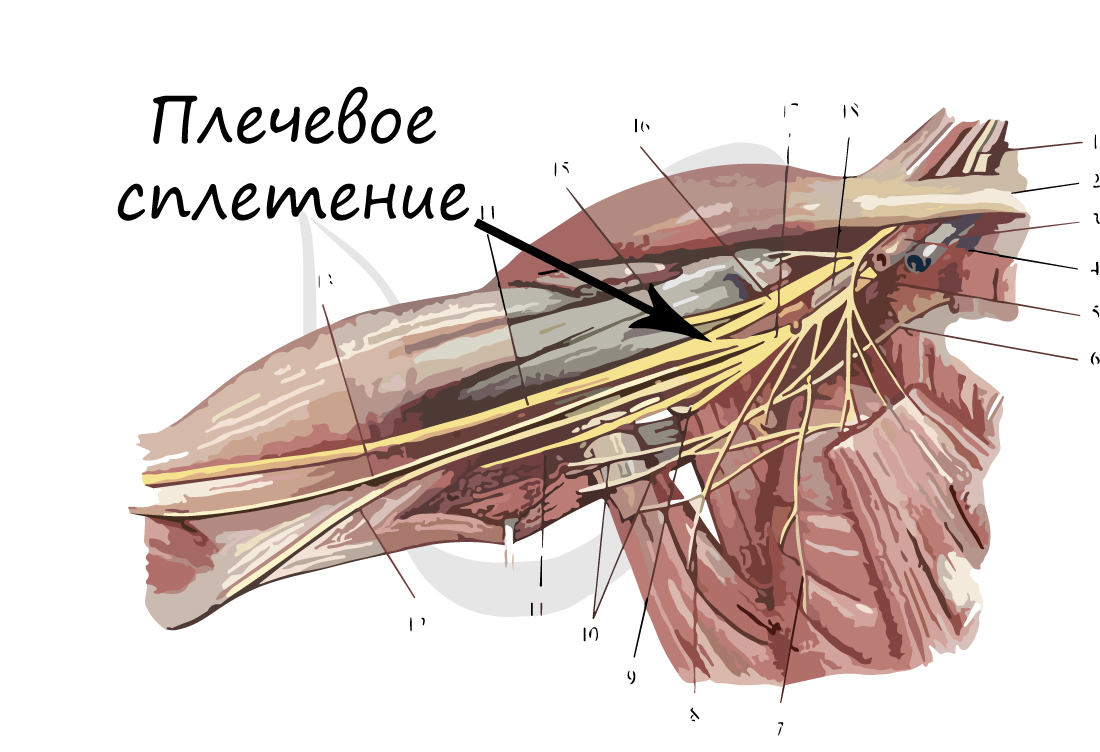
Нервы и нервные узлы
Собираясь вместе, отростки нейронов (нервные волокна) образуют пучки нервных волокон. Нервные пучки объединяются в нервы, которые покрыты соединительнотканной оболочкой.
В случае, если тела нейронов концентрируются в одном месте за пределами центральной нервной системы, их скопления
называют нервным узлом — или ганглием (от др.-греч. γάγγλιον — узел).
В случае сложных соединений между нервными волокнами говорят о нервных сплетениях. Одно из наиболее известных —
плечевое сплетение.
Болезни нервной системы
Неврологические болезни могут развиваться в любой точке нервной системы: от этого будет зависеть клиническая картина. В случае повреждения
чувствительного пути пациент перестает чувствовать боль, холод, тепло и другие раздражители в зоне иннервации пораженного нерва, при этом
движения сохранены в полном объеме.
Если повреждено двигательное звено, движение в пораженной конечности будет
невозможно: возникает паралич, но чувствительность может сохраняться.
Существует тяжелое мышечное заболеванием — миастения (от др.-греч. μῦς — «мышца» и ἀσθένεια — «бессилие, слабость»), при
котором собственные антитела разрушают мотонейроны (двигательные нейроны).

Постепенно любые движения мышцами становятся для пациента все труднее,
становится тяжело долго говорить, повышается утомляемость. Наблюдается характерный симптом — опущение верхнего века.
Болезнь может привести к слабости диафрагмы и дыхательных мышц, вследствие чего дыхание становится невозможным.
© Беллевич Юрий Сергеевич 2018-2023
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
| Моторный нейрон | |
|---|---|
 Микрофотография из подъязычного ядра, показывая двигательные нейроны с их характерными грубым Нисслит вещество ( «тигроид» цитоплазма). Окраска H amp; E-LFB. Микрофотография из подъязычного ядра, показывая двигательные нейроны с их характерными грубым Нисслит вещество ( «тигроид» цитоплазма). Окраска H amp; E-LFB. |
|
| Подробности | |
| Место нахождения | Брюшной рог из спинного мозга, некоторые черепные ядра нерва |
| Форма | Проекционный нейрон |
| Функция | Возбуждающая проекция (в НМС ) |
| Нейротрансмиттер | UMN в LMN : глутамат ; ЛМН к НМС : АХ |
| Пресинаптические связи | Первичная моторная кора через кортикоспинальный тракт |
| Постсинаптические связи | Мышечные волокна и другие нейроны |
| Идентификаторы | |
| MeSH | D009046 |
| НейроЛекс ID | nifext_103 |
| TA98 | A14.2.00.021 |
| TA2 | 6131 |
| FMA | 83617 |
| Анатомические термины нейроанатомии [ редактировать в Викиданных ] |
Двигательный нейрон (или мотонейронов или эфферентных нейрон) является нейрон, чьи клетки тела находится в моторной коре, стволе головного мозга или спинного мозга, и чьи аксонов (оптоволоконные) проектов в спинном мозге или снаружи спинного мозга прямо или косвенно контролируют эффекторные органы, в основном мышцы и железы. Есть два типа мотонейронов — верхние мотонейроны и нижние мотонейроны. Аксоны из синапсов верхних мотонейронов на интернейроны в спинном мозге и иногда прямо на нижние мотонейроны. Аксоны нижних мотонейронов представляют собой эфферентные нервные волокна, передающие сигналы от спинного мозга к эффекторам. Типы нижних мотонейронов — это альфа-мотонейроны, бета-мотонейроны и гамма-мотонейроны.
Один двигательный нейрон может иннервировать множество мышечных волокон, а мышечное волокно может подвергаться множеству потенциалов действия за время, необходимое для одного мышечного подергивания. Иннервация происходит в нервно-мышечном соединении, и судороги могут накладываться друг на друга в результате суммирования или тетанического сокращения. Отдельные подергивания могут стать неразличимыми, а напряжение плавно нарастает, в конечном итоге достигая плато.
СОДЕРЖАНИЕ
- 1 Развитие
- 2 Анатомия и физиология
- 2.1 Верхние двигательные нейроны
- 2.2 Нервные пути
- 2.3 Нижние двигательные нейроны
- 2.3.1 Соматические мотонейроны
- 2.3.2 Специальные висцеральные двигательные нейроны
- 2.3.3 Общие висцеральные двигательные нейроны
- 2.4 Нервно-мышечные соединения
- 3 См. Также
- 4 ссылки
- 5 Источники
Разработка
Моторные нейроны начинают развиваться на ранних этапах эмбрионального развития, и двигательные функции продолжают развиваться и в детстве. В нервной трубке клетки определяются либо по рострально-каудальной оси, либо по вентрально-дорсальной оси. В аксоны двигательных нейронов начинают появляться на четвертой неделе развития из вентральной области вентральной-спинные оси ( донца ). Этот гомеодомен известен как домен предшественников моторных нейронов (pMN). Факторы транскрипции здесь включают Pax6, OLIG2, Nkx-6.1 и Nkx-6.2, которые регулируются sonic hedgehog (Shh). Ген OLIG2 является наиболее важным из-за его роли в стимулировании экспрессии Ngn2, гена, который вызывает выход из клеточного цикла, а также способствует дальнейшим факторам транскрипции, связанным с развитием моторных нейронов.
Дальнейшая спецификация мотонейронов происходит, когда ретиноевая кислота, фактор роста фибробластов, Wnts и TGFb интегрируются в различные факторы транскрипции Hox. Существует 13 факторов транскрипции Hox, и вместе с сигналами они определяют, будет ли мотонейрон более ростральным или каудальным по характеру. В позвоночнике Hox 4-11 сортируют мотонейроны по одному из пяти моторных столбцов.
| Моторная колонка | Расположение в спинном мозге | Цель |
| Срединная моторная колонка | Присутствует по всей длине | Осевые мышцы |
| Гипаксиальная моторная колонка | Грудной отдел | Мышцы стенки тела |
| Преганглионарная моторная колонка | Грудной отдел | Симпатический ганглий |
| Боковая моторная колонка | Плечевая и поясничная области (обе области далее делятся на медиальную и латеральную области) | Мышцы конечностей |
| Диафрагмальная моторная колонка | Шейный отдел | Диафрагма |
Анатомия и психология
 Тракты спинного мозга
Тракты спинного мозга  Расположение нижних мотонейронов в спинном мозге
Расположение нижних мотонейронов в спинном мозге
Верхние двигательные нейроны
Верхние мотонейроны берут начало в моторной коре, расположенной в прецентральной извилине. Клетки, составляющие первичную моторную кору, — это клетки Беца, которые представляют собой тип пирамидных клеток. Аксоны этих клеток спускаются из коры, образуя кортикоспинальный тракт. Кортико- моторные нейроны проецируются из первичной коры прямо на мотонейроны вентрального рога спинного мозга. Их аксоны синапсы на спинномозговых мотонейронах множества мышц, а также на спинномозговых интернейронах. Они уникальны для приматов, и было высказано предположение, что их функцией является адаптивное управление руками, включая относительно независимое управление отдельными пальцами. Кортикомоторнейроны до сих пор были обнаружены только в первичной моторной коре, но не во вторичных моторных областях.
Нервные тракты
Нервные тракты представляют собой пучки аксонов в виде белого вещества, которые переносят потенциалы действия к своим эффекторам. В спинном мозге по этим нисходящим путям проходят импульсы из разных регионов. Эти тракты также служат местом происхождения нижних мотонейронов. В спинном мозге можно обнаружить семь основных нисходящих моторных трактов:
- Боковой кортикоспинальный тракт
- Руброспинальный тракт
- Боковой ретикулоспинальный тракт
- Вестибулоспинальный тракт
- Медиальный ретикулоспинальный тракт
- Тектоспинальный тракт
- Передний кортикоспинальный тракт
Нижние двигательные нейроны
Нижние мотонейроны — это те, которые берут начало в спинном мозге и прямо или косвенно иннервируют эффекторные мишени. Мишени этих нейронов различны, но в соматической нервной системе целью будет какое-то мышечное волокно. Есть три основных категории низших мотонейронов, которые можно разделить на подкатегории.
В соответствии с их целями мотонейроны подразделяются на три широкие категории:
- Соматические двигательные нейроны
- Специальные висцеральные двигательные нейроны
- Общие висцеральные двигательные нейроны
Соматические двигательные нейроны
Соматические двигательные нейроны берут начало в центральной нервной системе, проецируют свои аксоны на скелетные мышцы (например, мышцы конечностей, брюшные и межреберные мышцы ), которые участвуют в передвижении. Три типа этих нейронов — это альфа-эфферентные нейроны, бета-эфферентные нейроны и гамма-эфферентные нейроны. Они называются эфферентными, чтобы указывать на поток информации от центральной нервной системы (ЦНС) к периферии.
- Альфа-мотонейроны иннервируют экстрафузальные мышечные волокна, которые являются основным компонентом мышцы, генерирующим силу. Их клеточные тела находятся в вентральном роге спинного мозга, и их иногда называют клетками вентрального рога. Один мотонейрон может синапсировать в среднем со 150 мышечными волокнами. Моторный нейрон и все мышечные волокна, с которыми он соединяется, представляют собой двигательную единицу. Моторные агрегаты делятся на 3 категории: Основная статья: Моторные агрегаты.
- Медленные (S) двигательные единицы стимулируют мелкие мышечные волокна, которые сокращаются очень медленно и дают небольшое количество энергии, но очень устойчивы к утомлению, поэтому они используются для поддержания мышечного сокращения, например, для удержания тела в вертикальном положении. Они получают энергию с помощью окислительных средств и, следовательно, нуждаются в кислороде. Их еще называют красными волокнами.
- Двигательные единицы с быстрым утомлением (FF) стимулируют большие группы мышц, которые прилагают большие усилия, но очень быстро утомляются. Они используются для задач, требующих больших коротких всплесков энергии, таких как прыжки или бег. Они получают энергию с помощью гликолитических средств и, следовательно, не нуждаются в кислороде. Их называют белыми волокнами.
- Двигательные единицы, устойчивые к быстрому утомлению, стимулируют группы мышц среднего размера, которые не реагируют так быстро, как двигательные единицы FF, но могут удерживаться намного дольше (как следует из названия) и обеспечивают большую силу, чем двигательные единицы S. Они используют как окислительные, так и гликолитические средства для получения энергии.
В дополнение к произвольному сокращению скелетных мышц, альфа-мотонейроны также вносят вклад в мышечный тонус — постоянную силу, создаваемую несокращающимися мышцами для противодействия растяжению. Когда мышца растягивается, сенсорные нейроны в мышечном веретене определяют степень растяжения и посылают сигнал в ЦНС. ЦНС активирует альфа-двигательные нейроны в спинном мозге, которые заставляют экстрафузальные мышечные волокна сокращаться и тем самым сопротивляться дальнейшему растяжению. Этот процесс также называют рефлексом растяжения.
- Бета — мотонейроны иннервируют интрафузальные мышечные волокна из мышечных веретен, с залогами до extrafusal волокон. Есть два типа бета-мотонейронов: Медленно сокращающиеся — они иннервируют экстрафузальные волокна. Быстрое сокращение — они иннервируют интрафузальные волокна.
- Гамма-мотонейроны иннервируют интрафузальные мышечные волокна, находящиеся в мышечном веретене. Они регулируют чувствительность веретена к растяжению мышц. При активации гамма-нейронов интрафузальные мышечные волокна сокращаются, так что для активации сенсорных нейронов веретена и рефлекса растяжения требуется лишь небольшое растяжение. Существует два типа гамма-мотонейронов: динамические — они фокусируются на волокнах Bag1 и повышают динамическую чувствительность. Статические — они сосредоточены на волокнах Bag2 и повышают чувствительность к растяжению.
- Регулирующие факторы нижних мотонейронов
- Принцип размера — это относится к соме моторного нейрона. Это ограничивает более крупные нейроны для получения большего возбуждающего сигнала для стимуляции мышечных волокон, которые они иннервируют. Уменьшая количество ненужных мышечных волокон, организм может оптимизировать потребление энергии.
- Постоянный внутренний ток (PIC) — недавнее исследование на животных показало, что постоянный поток ионов, таких как кальций и натрий, через каналы в соме и дендритах влияет на синаптический вход. Другой способ думать об этом состоит в том, что постсинаптический нейрон активируется перед получением импульса.
- После гиперполяризации (AHP) — была выявлена тенденция, согласно которой медленные двигательные нейроны имеют более интенсивные AHP в течение более длительного времени. Один из способов запомнить это — то, что медленные мышечные волокна могут сокращаться дольше, поэтому имеет смысл, чтобы соответствующие им двигательные нейроны активировались медленнее.
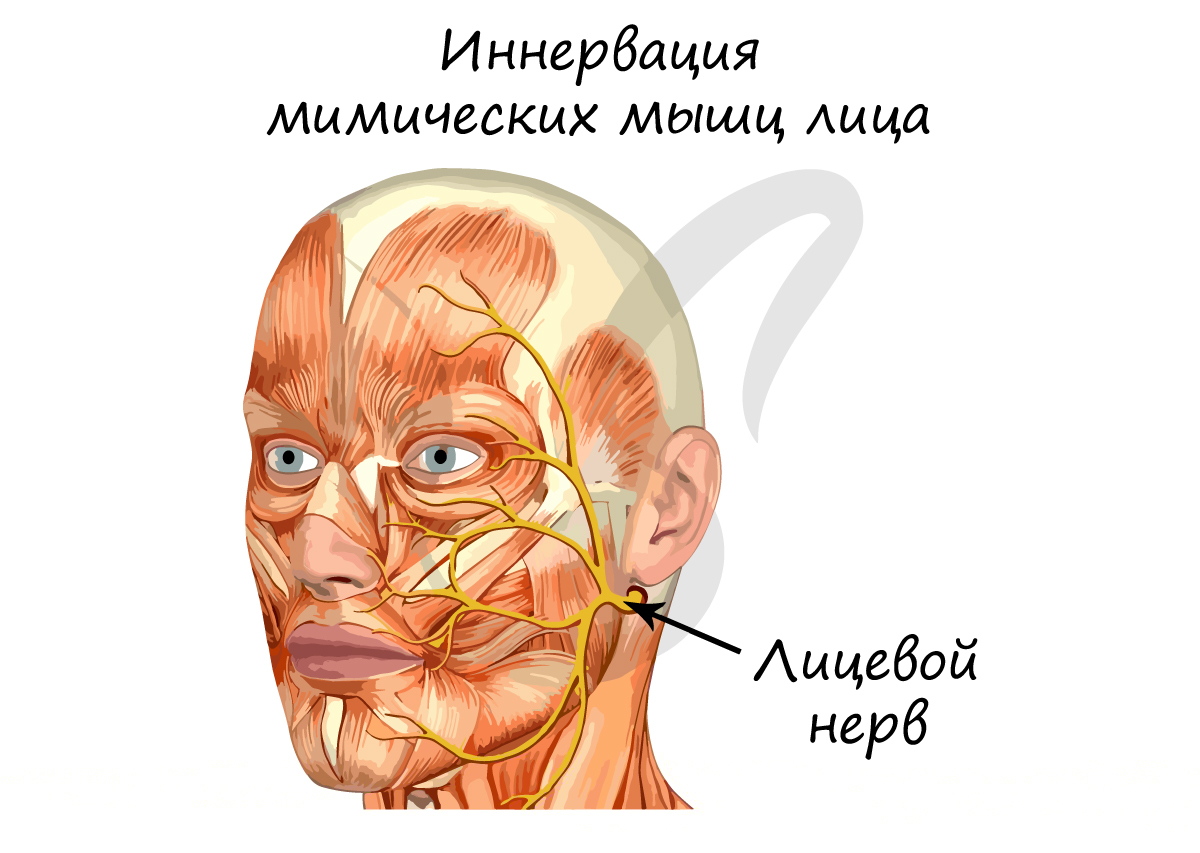
Специальные висцеральные двигательные нейроны
Они также известны как жаберные двигательные нейроны, которые участвуют в мимике, жевании, фонации и глотании. Связанные черепные нервы — глазодвигательный, отводящий, блокированный и подъязычный нервы.
| Филиал НС | Позиция | Нейротрансмиттер |
|---|---|---|
| Соматический | н / д | Ацетилхолин |
| Парасимпатический | Преганглионарный | Ацетилхолин |
| Парасимпатический | Ганглионарный | Ацетилхолин |
| Симпатичный | Преганглионарный | Ацетилхолин |
| Симпатичный | Ганглионарный | Норэпинефрин * |
| * За исключением волокон потовых желез и некоторых кровеносных сосудов, нейромедиаторов двигательных нейронов. |
Общие висцеральные двигательные нейроны
Эти моторные нейроны косвенно иннервируют сердечная мышца и гладких мышц на внутренностях (мышцы артерий ): они синапс на нейроны, расположенные в ганглиях от вегетативной нервной системы ( симпатической и парасимпатической ), расположенной в периферической нервной системе (ПНС), который сами непосредственно иннервируют висцеральные мышцы (а также некоторые клетки железы).
Как следствие, моторная команда скелетных и жаберных мышц является моносинаптической и включает только один мотонейрон, соматический или жаберный, который синапсирует с мышцами. Для сравнения, управление висцеральными мышцами является дисинаптическим с участием двух нейронов: общий висцеральный мотонейрон, расположенный в ЦНС, синапсирует с ганглиозным нейроном, расположенным в ПНС, который синапсирует с мышцей.
Все двигательные нейроны позвоночных являются холинэргическими, то есть они выделяют нейромедиатор ацетилхолин. Парасимпатические ганглиозные нейроны также являются холинергическими, тогда как большинство симпатических ганглиозных нейронов являются норадренергическими, то есть они выделяют нейромедиатор норадреналин. (см. таблицу)
Нервно-мышечные соединения
Один двигательный нейрон может иннервировать множество мышечных волокон, а мышечное волокно может подвергаться множеству потенциалов действия за время, необходимое для одного мышечного подергивания. В результате, если потенциал действия достигает до того, как судорога была завершена, подергивания могут накладываться друг на друг, либо через суммирование или тетаническое сжатие. В итоге мышца стимулируется повторно, так что дополнительные потенциалы действия, исходящие от соматической нервной системы, поступают до окончания подергивания. Таким образом, подергивания накладываются друг на друга, приводя к большей силе, чем при одиночном подергивании. Тетаническое сокращение вызывается постоянной, очень высокочастотной стимуляцией — потенциалы действия возникают с такой высокой скоростью, что отдельные подергивания неразличимы, а напряжение плавно нарастает, в конечном итоге достигая плато.
Интерфейс между двигательным нейроном и мышечным волокном представляет собой специализированный синапс, называемый нервно-мышечным соединением. При адекватной стимуляции мотонейрон высвобождает поток нейротрансмиттеров ацетилхолина (Ach) из окончаний аксона из синаптических везикул, связывающихся с плазматической мембраной. Молекулы ацетилхолина связываются с постсинаптическими рецепторами, находящимися внутри моторной концевой пластинки. После связывания двух рецепторов ацетилхолина открывается ионный канал, и ионы натрия могут проникать в клетку. Приток натрия в клетку вызывает деполяризацию и запускает мышечный потенциал действия. Затем стимулируются Т-канальцы сарколеммы, чтобы вызвать высвобождение ионов кальция из саркоплазматического ретикулума. Именно это химическое высвобождение заставляет целевое мышечное волокно сокращаться.
У беспозвоночных, в зависимости от высвобождаемого нейромедиатора и типа рецептора, который он связывает, реакция мышечного волокна может быть либо возбуждающей, либо тормозящей. Однако у позвоночных ответ мышечного волокна на нейромедиатор может быть только возбуждающим, другими словами, сократительным. Расслабление мышц и подавление мышечных сокращений у позвоночных достигаются только путем подавления самого двигательного нейрона. Именно так действуют миорелаксанты, воздействуя на двигательные нейроны, которые иннервируют мышцы (снижая их электрофизиологическую активность) или на холинергические нервно-мышечные соединения, а не на сами мышцы.
Смотрите также
- Ячейка Беца
- Центральный хроматолиз
- Двигательная дисфункция
- Заболевание двигательных нейронов
- Нерв
использованная литература
Источники
- Шервуд, Л. (2001). Физиология человека: от клеток к системам (4-е изд.). Пасифик Гроув, Калифорния: Брукс-Коул. ISBN 0-534-37254-6.
- Marieb, EN; Маллатт, Дж. (1997). Анатомия человека (2-е изд.). Менло-Парк, Калифорния: Бенджамин / Каммингс. ISBN 0-8053-4068-8.
From Wikipedia, the free encyclopedia
| Motor neurons | |
|---|---|
|
Micrograph of the hypoglossal nucleus showing motor neurons with their characteristic coarse Nissl substance («tigroid» cytoplasm). H&E-LFB stain. |
|
| Details | |
| Location | Ventral horn of the spinal cord, some cranial nerve nuclei |
| Shape | Projection neuron |
| Function | Excitatory projection (to NMJ) |
| Neurotransmitter | UMN to LMN: glutamate; LMN to NMJ: ACh |
| Presynaptic connections | Primary motor cortex via the Corticospinal tract |
| Postsynaptic connections | Muscle fibers and other neurons |
| Identifiers | |
| MeSH | D009046 |
| NeuroLex ID | nifext_103 |
| TA98 | A14.2.00.021 |
| TA2 | 6131 |
| FMA | 83617 |
| Anatomical terms of neuroanatomy
[edit on Wikidata] |
A motor neuron (or motoneuron or efferent neuron[1]) is a neuron whose cell body is located in the motor cortex, brainstem or the spinal cord, and whose axon (fiber) projects to the spinal cord or outside of the spinal cord to directly or indirectly control effector organs, mainly muscles and glands.[2] There are two types of motor neuron – upper motor neurons and lower motor neurons. Axons from upper motor neurons synapse onto interneurons in the spinal cord and occasionally directly onto lower motor neurons.[3] The axons from the lower motor neurons are efferent nerve fibers that carry signals from the spinal cord to the effectors.[4] Types of lower motor neurons are alpha motor neurons, beta motor neurons, and gamma motor neurons.
A single motor neuron may innervate many muscle fibres and a muscle fibre can undergo many action potentials in the time taken for a single muscle twitch. Innervation takes place at a neuromuscular junction and twitches can become superimposed as a result of summation or a tetanic contraction. Individual twitches can become indistinguishable, and tension rises smoothly eventually reaching a plateau.[5]
Development[edit]
Motor neurons begin to develop early in embryonic development, and motor function continues to develop well into childhood.[6] In the neural tube cells are specified to either the rostral-caudal axis or ventral-dorsal axis. The axons of motor neurons begin to appear in the fourth week of development from the ventral region of the ventral-dorsal axis (the basal plate).[7] This homeodomain is known as the motor neural progenitor domain (pMN). Transcription factors here include Pax6, OLIG2, Nkx-6.1, and Nkx-6.2, which are regulated by sonic hedgehog (Shh). The OLIG2 gene being the most important due to its role in promoting Ngn2 expression, a gene that causes cell cycle exiting as well as promoting further transcription factors associated with motor neuron development.[8]
Further specification of motor neurons occurs when retinoic acid, fibroblast growth factor, Wnts, and TGFb, are integrated into the various Hox transcription factors. There are 13 Hox transcription factors and along with the signals, determine whether a motor neuron will be more rostral or caudal in character. In the spinal column, Hox 4-11 sort motor neurons to one of the five motor columns.[8]
| Motor column | Location in spinal cord | Target |
| Median motor column | Present entire length | Axial muscles |
| Hypaxial motor column | Thoracic region | Body wall muscles |
| Preganglionic motor column | Thoracic region | Sympathetic ganglion |
| Lateral motor column | Brachial and lumbar region (both regions are further divided into medial and lateral domains) | Muscles of the limbs |
| Phrenic motor column | Cervical region | Diaphragm[10] |
Anatomy and physiology[edit]
Location of lower motor neurons in spinal cord
Upper motor neurons[edit]
Upper motor neurons originate in the motor cortex located in the precentral gyrus. The cells that make up the primary motor cortex are Betz cells, which are giant pyramidal cells. The axons of these cells descend from the cortex to form the corticospinal tract.[11] Corticomotorneurons project from the primary cortex directly onto motor neurons in the ventral horn of the spinal cord.[12][13] Their axons synapse on the spinal motor neurons of multiple muscles as well as on spinal interneurons.[12][13] They are unique to primates and it has been suggested that their function is the adaptive control of the hands including the relatively independent control of individual fingers.[13][14] Corticomotorneurons have so far only been found in the primary motor cortex and not in secondary motor areas.[13]
Nerve tracts[edit]
Nerve tracts are bundles of axons as white matter, that carry action potentials to their effectors. In the spinal cord these descending tracts carry impulses from different regions. These tracts also serve as the place of origin for lower motor neurons. There are seven major descending motor tracts to be found in the spinal cord:[15]
- Lateral corticospinal tract
- Rubrospinal tract
- Lateral reticulospinal tract
- Vestibulospinal tract
- Medial reticulospinal tract
- Tectospinal tract
- Anterior corticospinal tract
Lower motor neurons[edit]
Lower motor neurons are those that originate in the spinal cord and directly or indirectly innervate effector targets. The target of these neurons varies, but in the somatic nervous system the target will be some sort of muscle fiber. There are three primary categories of lower motor neurons, which can be further divided in sub-categories.[16]
According to their targets, motor neurons are classified into three broad categories:[17]
- Somatic motor neurons
- Special visceral motor neurons
- General visceral motor neurons
Somatic motor neurons[edit]
Somatic motor neurons originate in the central nervous system, project their axons to skeletal muscles[18] (such as the muscles of the limbs, abdominal, and intercostal muscles), which are involved in locomotion. The three types of these neurons are the alpha efferent neurons, beta efferent neurons, and gamma efferent neurons. They are called efferent to indicate the flow of information from the central nervous system (CNS) to the periphery.
- Alpha motor neurons innervate extrafusal muscle fibers, which are the main force-generating component of a muscle. Their cell bodies are in the ventral horn of the spinal cord and they are sometimes called ventral horn cells. A single motor neuron may synapse with 150 muscle fibers on average.[19] The motor neuron and all of the muscle fibers to which it connects is a motor unit. Motor units are split up into 3 categories:[20]
- Slow (S) motor units stimulate small muscle fibers, which contract very slowly and provide small amounts of energy but are very resistant to fatigue, so they are used to sustain muscular contraction, such as keeping the body upright. They gain their energy via oxidative means and hence require oxygen. They are also called red fibers.[20]
- Fast fatiguing (FF) motor units stimulate larger muscle groups, which apply large amounts of force but fatigue very quickly. They are used for tasks that require large brief bursts of energy, such as jumping or running. They gain their energy via glycolytic means and hence don’t require oxygen. They are called white fibers.[20]
- Fast fatigue-resistant motor units stimulate moderate-sized muscles groups that don’t react as fast as the FF motor units, but can be sustained much longer (as implied by the name) and provide more force than S motor units. These use both oxidative and glycolytic means to gain energy.[20]
In addition to voluntary skeletal muscle contraction, alpha motor neurons also contribute to muscle tone, the continuous force generated by noncontracting muscle to oppose stretching. When a muscle is stretched, sensory neurons within the muscle spindle detect the degree of stretch and send a signal to the CNS. The CNS activates alpha motor neurons in the spinal cord, which cause extrafusal muscle fibers to contract and thereby resist further stretching. This process is also called the stretch reflex.
- Beta motor neurons innervate intrafusal muscle fibers of muscle spindles, with collaterals to extrafusal fibres. There are two types of beta motor neurons: Slow Contracting- These innervate extrafusal fibers. Fast Contracting- These innervate intrafusal fibers.[21]
- Gamma motor neurons innervate intrafusal muscle fibers found within the muscle spindle. They regulate the sensitivity of the spindle to muscle stretching. With activation of gamma neurons, intrafusal muscle fibers contract so that only a small stretch is required to activate spindle sensory neurons and the stretch reflex. There are two types of gamma motor neurons: Dynamic- These focus on Bag1 fibers and enhance dynamic sensitivity. Static- These focus on Bag2 fibers and enhance stretch sensitivity.[21]
- Regulatory factors of lower motor neurons
- Size Principle – this relates to the soma of the motor neuron. This restricts larger neurons to receive a larger excitatory signal in order to stimulate the muscle fibers it innervates. By reducing unnecessary muscle fiber recruitment, the body is able to optimize energy consumption.[21]
- Persistent Inward Current (PIC) – recent animal study research has shown that constant flow of ions such as calcium and sodium through channels in the soma and dendrites influence the synaptic input. An alternate way to think of this is that the post-synaptic neuron is being primed before receiving an impulse.[21]
- After Hyper-polarization (AHP) – A trend has been identified that shows slow motor neurons to have more intense AHPs for a longer duration. One way to remember this is that slow muscle fibers can contract for longer, so it makes sense that their corresponding motor neurons fire at a slower rate.[21]
Special visceral motor neurons[edit]
These are also known as branchial motor neurons, which are involved in facial expression, mastication, phonation, and swallowing. Associated cranial nerves are the oculomotor, abducens, trochlear, and hypoglossal nerves.[17]
| Branch of NS | Position | Neurotransmitter |
|---|---|---|
| Somatic | n/a | Acetylcholine |
| Parasympathetic | Preganglionic | Acetylcholine |
| Parasympathetic | Ganglionic | Acetylcholine |
| Sympathetic | Preganglionic | Acetylcholine |
| Sympathetic | Ganglionic | Norepinephrine* |
| *Except fibers to sweat glands and certain blood vessels Motor neuron neurotransmitters |
General visceral motor neurons[edit]
These motor neurons indirectly innervate cardiac muscle and smooth muscles of the viscera ( the muscles of the arteries): they synapse onto neurons located in ganglia of the autonomic nervous system (sympathetic and parasympathetic), located in the peripheral nervous system (PNS), which themselves directly innervate visceral muscles (and also some gland cells).
In consequence, the motor command of skeletal and branchial muscles is monosynaptic involving only one motor neuron, either somatic or branchial, which synapses onto the muscle. Comparatively, the command of visceral muscles is disynaptic involving two neurons: the general visceral motor neuron, located in the CNS, synapses onto a ganglionic neuron, located in the PNS, which synapses onto the muscle.
All vertebrate motor neurons are cholinergic, that is, they release the neurotransmitter acetylcholine. Parasympathetic ganglionic neurons are also cholinergic, whereas most sympathetic ganglionic neurons are noradrenergic, that is, they release the neurotransmitter noradrenaline. (see Table)
Neuromuscular junctions[edit]
A single motor neuron may innervate many muscle fibres and a muscle fibre can undergo many action potentials in the time taken for a single muscle twitch. As a result, if an action potential arrives before a twitch has completed, the twitches can superimpose on one another, either through summation or a tetanic contraction. In summation, the muscle is stimulated repetitively such that additional action potentials coming from the somatic nervous system arrive before the end of the twitch. The twitches thus superimpose on one another, leading to a force greater than that of a single twitch. A tetanic contraction is caused by constant, very high frequency stimulation — the action potentials come at such a rapid rate that individual twitches are indistinguishable, and tension rises smoothly eventually reaching a plateau.[5]
The interface between a motor neuron and muscle fiber is a specialized synapse called the neuromuscular junction. Upon adequate stimulation, the motor neuron releases a flood of acetylcholine (Ach) neurotransmitters from the axon terminals from synaptic vesicles bind with the plasma membrane. The acetylcholine molecules bind to postsynaptic receptors found within the motor end plate. Once two acetylcholine receptors have been bound, an ion channel is opened and sodium ions are allowed to flow into the cell. The influx of sodium into the cell causes depolarization and triggers a muscle action potential. T tubules of the sarcolemma are then stimulated to elicit calcium ion release from the sarcoplasmic reticulum. It is this chemical release that causes the target muscle fiber to contract.[19]
In invertebrates, depending on the neurotransmitter released and the type of receptor it binds, the response in the muscle fiber could be either excitatory or inhibitory. For vertebrates, however, the response of a muscle fiber to a neurotransmitter can only be excitatory, in other words, contractile. Muscle relaxation and inhibition of muscle contraction in vertebrates is obtained only by inhibition of the motor neuron itself. This is how muscle relaxants work by acting on the motor neurons that innervate muscles (by decreasing their electrophysiological activity) or on cholinergic neuromuscular junctions, rather than on the muscles themselves.
Synaptic input to motor neurons[edit]
Motor neurons receive synaptic input from premotor neurons. Premotor neurons can be 1) spinal interneurons that have cell bodies in the spinal cord, 2) sensory neurons that convey information from the periphery and synapse directly onto motoneurons, 3) descending neurons that convey information from the brain and brainstem. The synapses can be excitatory, inhibitory, electrical, or neuromodulatory. For any given motor neuron, determining the relative contribution of different input sources is difficult, but advances in connectomics have made it possible for fruit fly motor neurons. In the fly, motor neurons controlling the legs and wings are found in the ventral nerve cord, homologous to the spinal cord. Fly motor neurons vary by over 100X in the total number of input synapses. However, each motor neuron gets similar fractions of its synapses from each premotor source: ~70% from neurons within the VNC, ~10% from descending neurons, ~3% from sensory neurons, and ~6% from VNC neurons that also send a process up to the brain. The remaining 10% of synapses come from neuronal fragments that are unidentified by current image segmentation algorithms and require additional manual segmentation to measure [22].
See also[edit]
- Betz cell
- Central chromatolysis
- Motor dysfunction
- Motor neuron disease
- Nerve
- Efferent nerve fiber
- Motor nerve
References[edit]
- ^ «Afferent vs. Efferent: AP® Psych Crash Course Review | Albert.io». Albert Resources. 2019-12-02. Retrieved 2021-04-25.
- ^ Tortora, Gerard; Derrickson, Bryan (2014). Principles of Anatomy & Physiology (14th ed.). New Jersey: John Wiley & Sons, Inc. pp. 406, 502, 541. ISBN 978-1-118-34500-9.
- ^ Pocock, Gillian; Richards, Christopher D. (2006). Human physiology : the basis of medicine (3rd ed.). Oxford: Oxford University Press. pp. 151–153. ISBN 978-0-19-856878-0.
- ^ Schacter D.L., Gilbert D.T., and Wegner D.M. (2011) Psychology second edition. New York, NY: Worth
- ^ a b Russell, Peter (2013). Biology — Exploring the Diversity of Life. Toronto: Nelson Education. p. 946. ISBN 978-0-17-665133-6.
- ^ Tortora, Gerard; Derrickson, Bryan (2011). Principles of Anatomy Physiology (14th ed.). New Jersey: John Wiley & Sons, Inc. pp. 1090–1099. ISBN 978-1-118-34500-9.
- ^ Sadler, T. (2010). Langman’s medical embryology (11th ed.). Philadelphia: Lippincott William & Wilkins. pp. 299–301. ISBN 978-0-7817-9069-7.
- ^ a b Davis-Dusenbery, BN; Williams, LA; Klim, JR; Eggan, K (February 2014). «How to make spinal motor neurons». Development. 141 (3): 491–501. doi:10.1242/dev.097410. PMID 24449832.
- ^ Edgar R, Mazor Y, Rinon A, Blumenthal J, Golan Y, Buzhor E, Livnat I, Ben-Ari S, Lieder I, Shitrit A, Gilboa Y, Ben-Yehudah A, Edri O, Shraga N, Bogoch Y, Leshansky L, Aharoni S, West MD, Warshawsky D, Shtrichman R (2013). «LifeMap Discovery™: The Embryonic Development, Stem Cells, and Regenerative Medicine Research Portal». PLOS ONE. 8 (7): e66629. Bibcode:2013PLoSO…866629E. doi:10.1371/journal.pone.0066629. ISSN 1932-6203. PMC 3714290. PMID 23874394.
- ^ Philippidou, Polyxeni; Walsh, Carolyn; Aubin, Josée; Jeannotte, Lucie; Dasen, Jeremy S. (2012). «Sustained Hox5 Gene Activity is Required for Respiratory Motor Neuron Development». Nature Neuroscience. 15 (12): 1636–1644. doi:10.1038/nn.3242. ISSN 1097-6256. PMC 3676175. PMID 23103965.
- ^ Fitzpatrick, D. (2001) The Primary Motor Cortex: Upper Motor Neurons That Initiate Complex Voluntary Movements. In D. Purves, G.J. Augustine, D. Fitzpatrick, et al. (Ed.), Neuroscience. Retrieved from «The Primary Motor Cortex: Upper Motor Neurons That Initiate Complex Voluntary Movements — Neuroscience — NCBI Bookshelf». Archived from the original on 2018-06-05. Retrieved 2017-11-30.
- ^ a b Mack, Sarah; Kandel, Eric R.; Jessell, Thomas M.; Schwartz, James H.; Siegelbaum, Steven A.; Hudspeth, A. J. (2013). Principles of neural science. Kandel, Eric R. (5th ed.). New York. ISBN 9780071390118. OCLC 795553723.
- ^ a b c d Lemon, Roger N. (April 4, 2008). «Descending Pathways in Motor Control». Annual Review of Neuroscience. 31 (1): 195–218. doi:10.1146/annurev.neuro.31.060407.125547. ISSN 0147-006X. PMID 18558853. S2CID 16139768.
- ^ Isa, T (April 2007). «Direct and indirect cortico-motoneuronal pathways and control of hand/arm movements». Physiology. 22 (2): 145–152. doi:10.1152/physiol.00045.2006. PMID 17420305.
- ^ Tortora, G. J., Derrickson, B. (2011). The Spinal Cord and Spinal Nerves. In B. Roesch, L. Elfers, K. Trost, et al. (Ed.), Principles of Anatomy and Physiology (pp. 443-468). New Jersey: John Wiley & Sons, Inc.
- ^ Fitzpatrick, D. (2001) Lower Motor Neuron Circuits and Motor Control: Overview. In D. Purves, G.J. Augustine, D. Fitzpatrick, et al. (Ed.), Neuroscience. Retrieved from «Lower Motor Neuron Circuits and Motor Control — Neuroscience — NCBI Bookshelf». Archived from the original on 2018-06-05. Retrieved 2017-11-30.
- ^ a b «CHAPTER NINE». www.unc.edu. Archived from the original on 2017-11-05. Retrieved 2017-12-08.
- ^ Silverthorn, Dee Unglaub (2010). Human Physiology: An Integrated Approach. Pearson. p. 398. ISBN 978-0-321-55980-7.
- ^ a b Tortora, G. J., Derrickson, B. (2011). Muscular Tissue. In B. Roesch, L. Elfers, K. Trost, et al. (Ed.), Principles of Anatomy and Physiology (pp. 305-307, 311). New Jersey: John Wiley & Sons, Inc.
- ^ a b c d Purves D, Augustine GJ, Fitzpatrick D, et al., editors: Neuroscience. 2nd edition, 2001 «The Motor Unit — Neuroscience — NCBI Bookshelf». Archived from the original on 2018-06-05. Retrieved 2017-09-05.
- ^ a b c d e Manuel, Marin; Zytnicki, Daniel (2011). «Alpha, Beta, and Gamma Motoneurons: Functional Diversity in the Motor System’s Final Pathway». Journal of Integrative Neuroscience. 10 (3): 243–276. doi:10.1142/S0219635211002786. ISSN 0219-6352. PMID 21960303. S2CID 21582283.
- ^ Azevedo, Anthony; Lesser, Ellen; Mark, Brandon; Phelps, Jasper; Elabbady, Leila; Kuroda, Sumiya; Sustar, Anne; Moussa, Anthony; Kandelwal, Avinash; Dallmann, Chris J.; Agrawal, Sweta; Lee, Su-Yee J.; Pratt, Brandon; Cook, Andrew; Skutt-Kakaria, Kyobi (2022-12-15). «Tools for comprehensive reconstruction and analysis of Drosophila motor circuits»: 2022.12.15.520299. doi:10.1101/2022.12.15.520299.
Sources[edit]
- Sherwood, L. (2001). Human Physiology: From Cells to Systems (4th ed.). Pacific Grove, CA: Brooks-Cole. ISBN 0-534-37254-6.
- Marieb, E. N.; Mallatt, J. (1997). Human Anatomy (2nd ed.). Menlo Park, CA: Benjamin/Cummings. ISBN 0-8053-4068-8.
Значение слова «мотонейрон»
Мотонейро́н (лат. motor — приводящий в движение + нейрон; син. двигательный нейрон) — крупная нервная клетка в передних рогах спинного мозга. Мотонейроны обеспечивают моторную координацию и поддержание мышечного тонуса.
Все значения слова «мотонейрон»
Предложения со словом «мотонейрон»
-
Управляющие сигналы из коры головного мозга направляются по нисходящим нервным путям к мотонейронам спинного мозга.
-
Аксоны клеток, образующих центры, переходят в передний рог и покидают спинной мозг в составе передних корешков вместе с аксонами и мотонейронами передних рогов.
-
Двигательные корешки содержат аксоны мотонейронов нейроцитов переднего рога серого вещества.
- (все предложения)
Правописание
- Как правильно пишется слово «мотонейрон»