8
Лекция 13
Биоэнергетика мышечной ткани
Как уже указывалось
ранее, обе фазы мышечной деятельности
– сокращение и расслабление — протекают
при обязательном использовании энергии,
которая выделяется при гидролизе АТФ:
АТФ +H2O
= АДФ + H3PO4
– ΔG
Однако запасы АТФ
в клетках мышечной ткани незначительны
(в покое концентрация АТФ составляет 5
ммоль/л) и их достаточно для мышечной
работы в течение 1-2 секунд. В связи с
этим для обеспечения более продолжительной
мышечной деятельности в мышцах должно
происходить пополнение запасов АТФ.
!!! Образование
АТФ непосредственно в мышечных клетках
во время физической работы называется
ресинтезом
АТФ и
идет с потреблением энергии.
В зависимости от
источника энергии выделяют несколько
путей расинтеза АТФ.
Источники
энергии следующие.
-
Окислительное
фосфорилирование. -
Гликолиз,
гликогенолиз. -
Специальные
реакции субстратного фосфорилирования.
13.1. Окислительное фосфорилирование.
Синонимы:
тканевое дыхание, аэробное или
окислительное фосфорилирование.
В
процессе окислительного фосфорилирования
от окисляемого субстрата отнимаются
два атома водорода (два протона и два
электрона) и по ЭТЦ передаются на
молекулярный кислород, доставляемый
кровью в мышцы из воздуха, в результате
чего образуется ПВК.
Важным
компонентом окислительного фосфорилирования
является цикл трикарбоновых кислот
(цикл Кребса, цикл лимонной кислоты):
2
изолимонная кислота →
2 α-кетоглутаровая кислота →
2 янтарная кислота →
2 яблочная
кислота
В
ходе перечисленного процесса от
перечисленных выше кислот отнимается
8 (2x4)
пары атомов водорода и поэтому образуется
24 молекулы АТФ. Совокупный энергетический
баланс окислительного фосфорилирования
(аэробный гликолиз + активация ПВК + ЦТК)
составляет 38 молекул АТФ.
Преимущества:
1.
Высокая экономичность – в ходе этого
процесса идёт глубокая распад окисляемых
веществ до конечных продуктов – CO2
H2O
и поэтому выделяется большое количество
энергии. Так, например, при аэробном
окислении мышечного гликогена образуется
38 молекул АТФ. в расчёте на каждую
отщепляемую от гликогена молекулу
глюкозы (при анаэробном гликолизе – 3
молекулы АТФ).
2.
Универсальность использования субстратов.
В ходе аэробного ресинтеза АТФ окисляются
все основные органические вещества
организма: аминокислоты, жирные кислоты,
углеводы, кетоновые тела и т.д.
3.
Продукты
распада (CO2
и H2O)
практически безвредны.
Недостатки:
1.
Требует повышенных количеств кислорода,
доставка которого в мышцы, обеспечивается
дыхательной сердечно-сосудистой
системами. Функциональное состояние
этих систем является лимитирующим
фактором, ограничивающим продолжительность
аэробного пути ресинтеза АТФ с максимальной
скоростью и виличину самой максимальной
мощности.
Важную роль в
обеспечении мышечной клетки кислородом
играет миоглобин, у которого сродство
к кислороду больше, чем у гемоглобина:
при парциальном давлении кислорода,
равном 30 мм.рт.ст., миоглобин насыщается
кислородом на 100 %, а гемоглобин — всего
на 30 %. Поэтому миоглобин эффективно
отнимает у гемоглобин доставляемый им
кислород.
2. Длительное время
развёртывания (3-4 минуты) и небольшую
по абсолютной величине максимальную
мощность.
В связи с этим,
интенсивная мышечная деятельность,
свойственная большинству видов спорта,
не может быть полностью обеспечена этим
путём ресинтеза АТФ. Мышцы вынуждены
дополнительно включать анаэробные
реакции субстратного фосфорилирования,
имеющие более короткое время развёртывания
и большую максимальную мощность.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Карта слов и выражений русского языка
Онлайн-тезаурус с возможностью поиска ассоциаций, синонимов, контекстных связей
и
примеров
предложений к словам и выражениям русского языка.
Справочная информация по склонению имён существительных и прилагательных,
спряжению
глаголов, а также
морфемному строению слов.
Сайт оснащён мощной системой поиска с
поддержкой русской морфологии.
Разбор слова
по составу ОНЛАЙН
Подобрать синонимы
ОНЛАЙН
Найти предложения со словом
или
выражением ОНЛАЙН
Поиск по произведениям русской классики
ОНЛАЙН
Словарь афоризмов русских писателей
На букву О Со слова «окислительное»
Фраза «окислительное фосфорилирование»
Фраза состоит из двух слов и 29 букв без пробелов.
- Синонимы к фразе
- Написание фразы наоборот
- Написание фразы в транслите
- Написание фразы шрифтом Брайля
- Передача фразы на азбуке Морзе
- Произношение фразы на дактильной азбуке
- Остальные фразы со слова «окислительное»
- Остальные фразы из 2 слов
 16:46
16:46
Аэробный этап клеточного дыхания. Окислительное фосфорилирование. Центр онлайн-обучения «Фоксфорд»
 05:16
05:16
Дыхательная цепь
 12:10
12:10
Цитология. Лекция 54. Окислительное фосфорилирование
 07:35
07:35
Дыхательная цепь (цепь транспорта электронов) митохондрии на русском (Electron Transport Chain).
 12:13
12:13
Окислительное фосфорилирование (видео 10) | Клеточное дыхание | Биология
 05:03
05:03
Окислительное фосфорилирование и хемиосмос (видео 9) | Клеточное дыхание | Биология
Синонимы к фразе «окислительное фосфорилирование»
Какие близкие по смыслу слова и фразы, а также похожие выражения существуют. Как можно написать по-другому или сказать другими словами.
Фразы
- + азотистый баланс −
- + активные формы кислорода −
- + биогенные амины −
- + биосинтез белка −
- + внутренняя мембрана митохондрий −
- + гормоны передней доли гипофиза −
- + градиент концентрации −
- + дыхательная цепь −
- + заменимые аминокислоты −
- + избирательная проницаемость −
- + карбоксильная группа −
- + кетоновые тела −
- + кислотный остаток −
- + костный матрикс −
- + мембранный потенциал −
- + мышечные клетки −
- + обратное всасывание −
- + окислительное фосфорилирование −
- + окислительные процессы −
- + окислительный стресс −
- + пассивный транспорт −
- + первичная моча −
- + перенос электрона −
- + плазма крови −
Ваш синоним добавлен!
Написание фразы «окислительное фосфорилирование» наоборот
Как эта фраза пишется в обратной последовательности.
еинаворилирофсоф еоньлетилсико 😀
Написание фразы «окислительное фосфорилирование» в транслите
Как эта фраза пишется в транслитерации.
в армянской🇦🇲 ոկիսլիտելնոե ֆոսֆորիլիրովանիե
в греческой🇬🇷 οκεισλειθαιλνοαι φοσφορειλειρουανειαι
в грузинской🇬🇪 ოკისლითელნოე ფოსფორილიროვანიე
в еврейской🇮🇱 וכיסליטאלנוא פוספורילירובאניא
в латинской🇬🇧 okislitelnoye fosforilirovaniye
Как эта фраза пишется в пьюникоде — Punycode, ACE-последовательность IDN
xn--e1aahaihcqch1ao6k xn--80adkjacrtdadudi7bc
Как эта фраза пишется в английской Qwerty-раскладке клавиатуры.
jrbckbntkmyjtajcajhbkbhjdfybt
Написание фразы «окислительное фосфорилирование» шрифтом Брайля
Как эта фраза пишется рельефно-точечным тактильным шрифтом.
⠕⠅⠊⠎⠇⠊⠞⠑⠇⠾⠝⠕⠑⠀⠋⠕⠎⠋⠕⠗⠊⠇⠊⠗⠕⠺⠁⠝⠊⠑
Передача фразы «окислительное фосфорилирование» на азбуке Морзе
Как эта фраза передаётся на морзянке.
– – – – ⋅ – ⋅ ⋅ ⋅ ⋅ ⋅ ⋅ – ⋅ ⋅ ⋅ ⋅ – ⋅ ⋅ – ⋅ ⋅ – ⋅ ⋅ – – ⋅ – – – ⋅ ⋅ ⋅ – ⋅ – – – ⋅ ⋅ ⋅ ⋅ ⋅ – ⋅ – – – ⋅ – ⋅ ⋅ ⋅ ⋅ – ⋅ ⋅ ⋅ ⋅ ⋅ – ⋅ – – – ⋅ – – ⋅ – – ⋅ ⋅ ⋅ ⋅
Произношение фразы «окислительное фосфорилирование» на дактильной азбуке
Как эта фраза произносится на ручной азбуке глухонемых (но не на языке жестов).
Передача фразы «окислительное фосфорилирование» семафорной азбукой
Как эта фраза передаётся флажковой сигнализацией.













Остальные фразы со слова «окислительное»
Какие ещё фразы начинаются с этого слова.
- окислительное дезаминирование
- окислительное декарбоксилирование пирувата
Ваша фраза добавлена!
Остальные фразы из 2 слов
Какие ещё фразы состоят из такого же количества слов.
- а вдобавок
- а вдруг
- а ведь
- а вот
- а если
- а ещё
- а именно
- а капелла
- а каторга
- а ну-ка
- а приятно
- а также
- а там
- а то
- аа говорит
- аа отвечает
- аа рассказывает
- ааронов жезл
- аароново благословение
- аароново согласие
- аб ово
- абажур лампы
- абазинская аристократия
- абазинская литература
Комментарии
 @gbgr 02.01.2020
@gbgr 02.01.2020 08:31
Что значит фраза «окислительное фосфорилирование»? Как это понять?..
Ответить
@eugbnno 13.09.2022 21:40
![]() 1
1
×
![]()
Здравствуйте!
У вас есть вопрос или вам нужна помощь?
![]()
Спасибо, ваш вопрос принят.
Ответ на него появится на сайте в ближайшее время.
А Б В Г Д Е Ё Ж З И Й К Л М Н О П Р С Т У Ф Х Ц Ч Ш Щ Ъ Ы Ь Э Ю Я
Транслит Пьюникод Шрифт Брайля Азбука Морзе Дактильная азбука Семафорная азбука
Палиндромы Сантана
Народный словарь великого и могучего живого великорусского языка.
Онлайн-словарь слов и выражений русского языка. Ассоциации к словам, синонимы слов, сочетаемость фраз. Морфологический разбор: склонение существительных и прилагательных, а также спряжение глаголов. Морфемный разбор по составу словоформ.
По всем вопросам просьба обращаться в письмошную.
Эта тема входит в мой курс по биохимии
Окислительное фосфорилирование — это заключительный этап общего катаболизма. Напомню, что он включает: окислительное декарбоксилирование пирувата, цикл Кребса и окислительное фосфорилирование. Про первые два говорили здесь.
Окислительное фосфорилирование — сложный процесс. Он включает три этапа: перенос электронов по дыхательной цепи, создание градиента концентрации протонов и синтез АТФ. Не слишком понятно, не правда ли? Ничего, сейчас разберёмся. Но сначала нужно вспомнить кое-что.
Общий путь катаболизма идёт в митохондрии. Нам нужно разобраться со строением мембран митохондрии, потому что без этого мы далеко не уедем. У митохондрии есть две мембраны: наружная и внутренняя. В наружной мембране есть поры, поэтому она проницаема для небольших молекул, в том числе ионов. Внутренняя мембрана непроницаема для любых молекул — там нет пор. Тут я загнул, всё-таки есть жирорастворимые молекулы, которым на мембрану плевать.
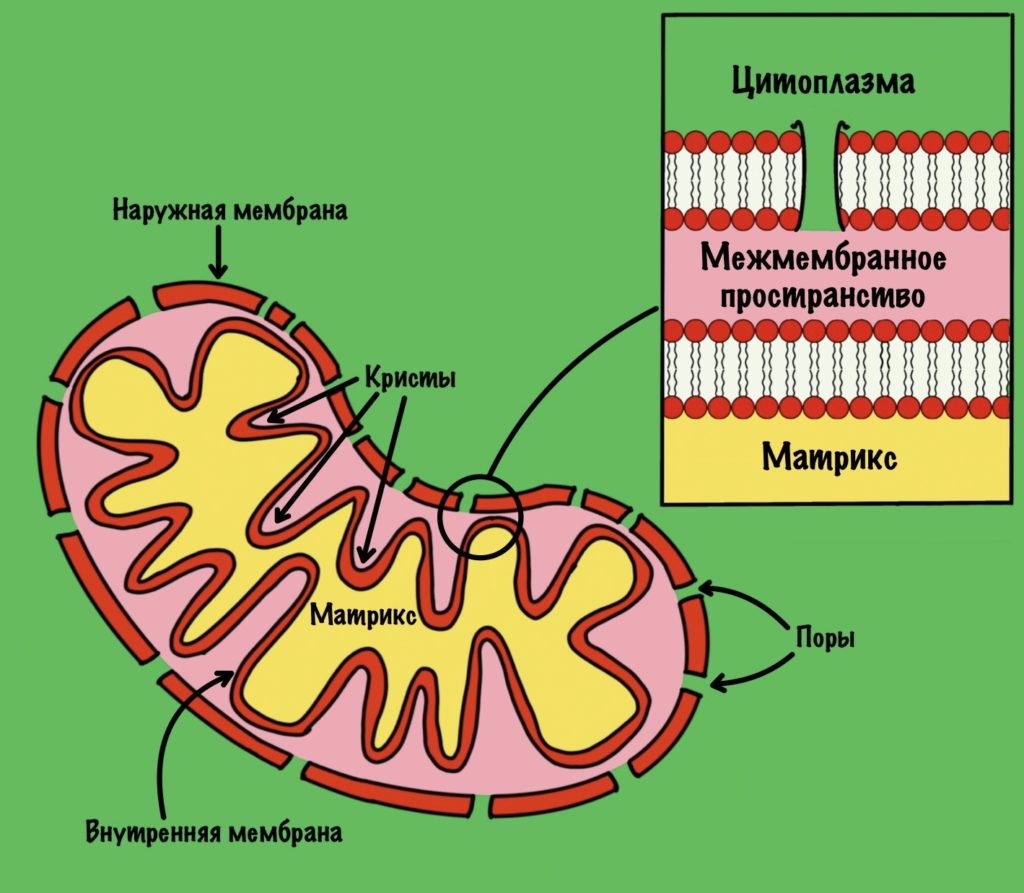
Внутренняя мембрана увеличивает свою площадь за счет формирования крист. Видите, какая она неровная? Матрикс — это пространство между внутренней мембраной. А между внутренней и наружной мембранами находится мемжмембранное пространство.
Зачем нужно было это вспомнить? Потому что во внутренней мембране находится дыхательная цепь (или цепь переноса электронов). Чем больше площадь этой мембраны, тем больше в ней комплексов дыхательной цепи. Да-да, она состоит из нескольких комплексов, но пока что нужно запомнить вот это:
Внутренняя мембрана непроницаема для молекул
Можно обойти этот запрет. Для этого нужны белки переносчики, которые перекинут молекулу на другую сторону.
Окислительное фосфорилирование
Теперь займёмся окислительным фосфорилированием. Какая была основная суть у катаболизма? Вытащить максимальное количество электронов из молекулы — с этим он отлично справился, если вспомнить катаболизм одной молекулы глюкозы, то в ходе неё были получены 24 электрона — в виде НАДH и ФАДH2. Из них: четыре в гликолизе, четыре в окислительном декарбоксилировании пирувата и шестнадцать в цикле трикарбоновых кислот.
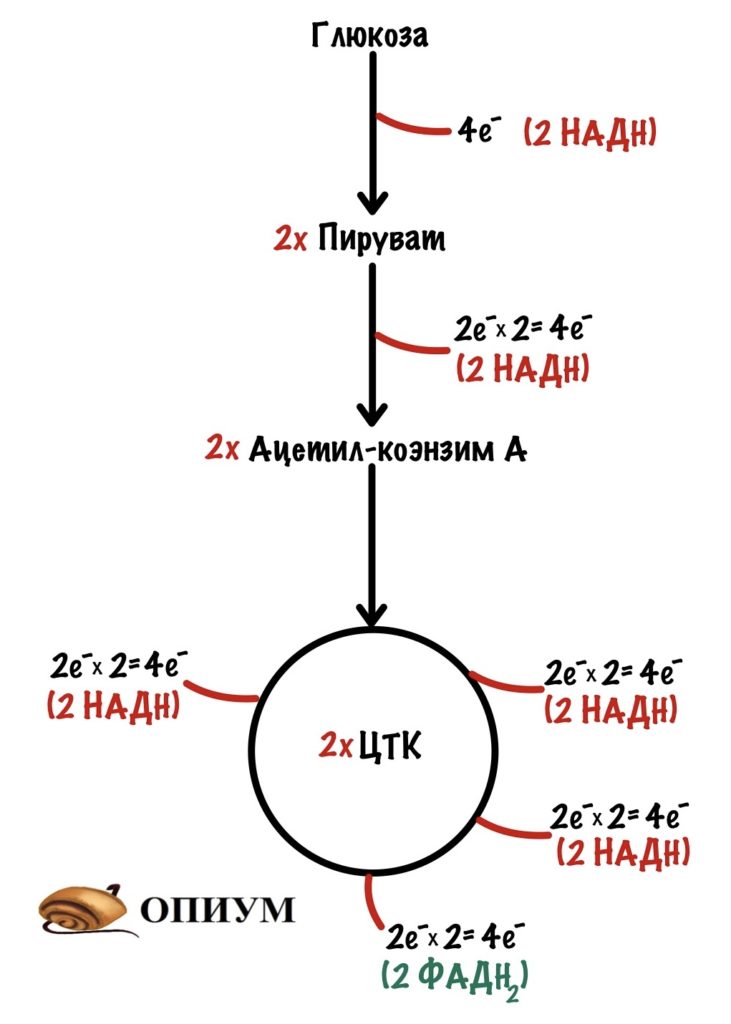
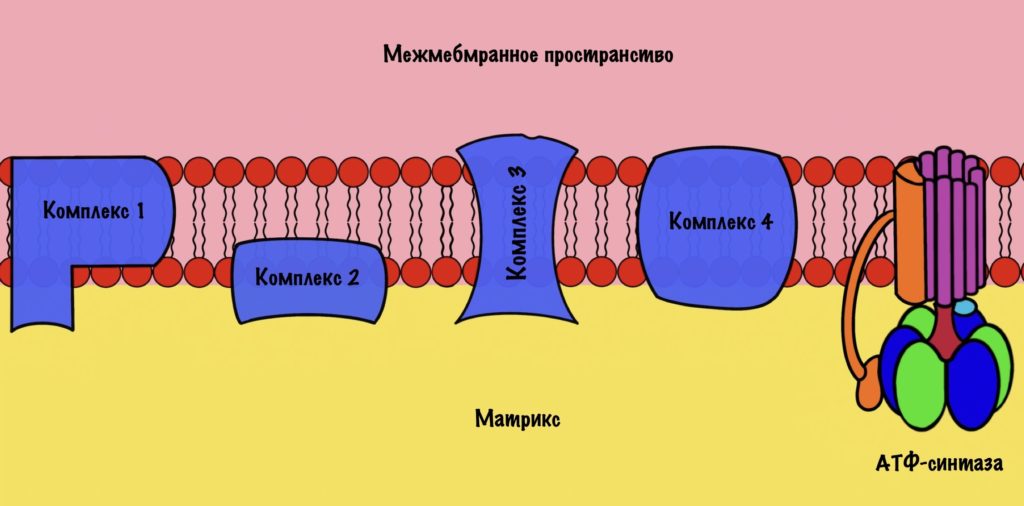
Зачем это было нужно? Для окислительного фосфорилирования, а если точнее, то одной из его частей — цепи переноса электронов. Мы договорились, что она находится во внутренней мембране митохондрий. Дыхательная цепь состоит из четырёх комплексов, но пока не будем лезть в дебри. Сейчас просто разберёмся с механизмом окислительного фофорилирования.
Электроны, которые были получены в ходе катаболизма, попадают на комплексы дыхательной цепи. Там они перескакивают с одного комплекса на другой, пока не попадут на кислород. В ходе таких перескоков создаётся энергия. Сразу скажу, что её не слишком много. Но достаточно для того, чтобы транспортировать протоны из матрикса в межмембранное пространство. За счёт этого создаётся градиент концентрации протонов. Протоны стараются попасть обратно в матрикс митохондрий, но мембрана для них непроницаема. Так они и бьются в мембрану, пока не попадают на АТФ-синтазу. Можно подумать, что АТФ-синтаза это канал для протонов, но это не совсем так. Транспорт протонов из межмембранного пространства в матрикс создаёт большое количество энергии. Эта энергия используется АТФ-синтазой для синтеза АТФ. Чем-то напоминает вторично-активный транспорт.
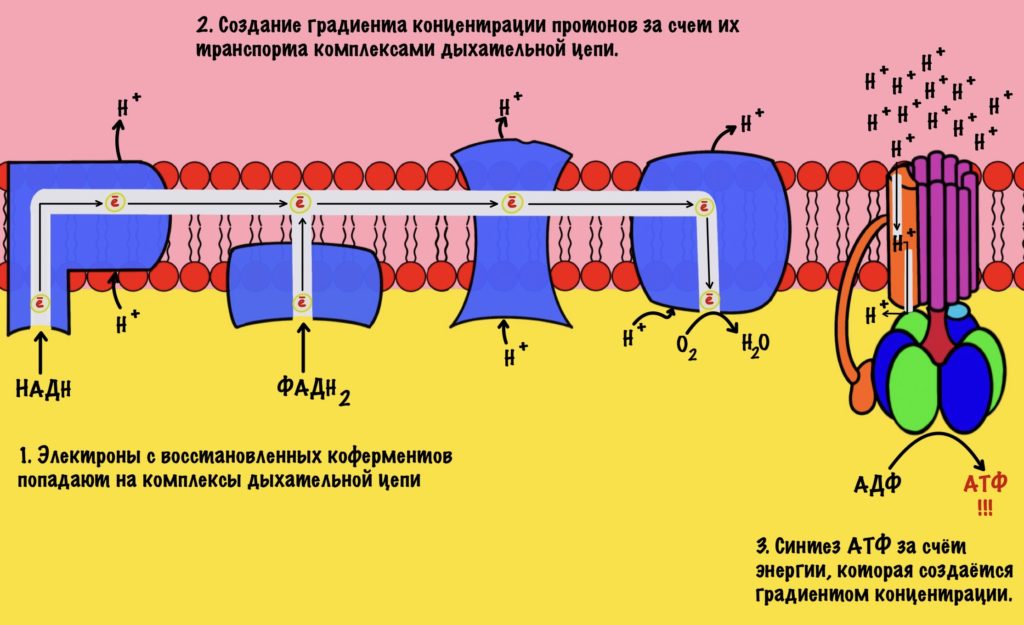
Представьте себе шланг. Вода под большим давлением идёт по шлангу, пока не дойдёт до его конца. Чем больше давление в шланге, тем сильнее из него вырывается вода. Примерно такой же принцип в работе окислительного фосфорилирования. Только напор воды создаёт дыхательная цепь, а единственное место выхода для неё — это АТФ-синтаза. Вода — это протоны водорода.
Из этого можно сделать вывод. Если мы не создадим градиент концентрации, то синтеза АТФ не будет. Получается, что работа цепи переноса электронов сопряжена с работой АТФ-синтазы. Это и есть окислительное фосфорилирование. В нём сопряжены три процесса: транспорт электронов по дыхательной цепи, транспорт протонов в межмембранное пространство и синтез АТФ. За первые два процесса отвечает дыхательная цепь, а за последний — АТФ-синтаза. Теперь, когда мы уяснили себе этот принцип, можно лезть в дебри!
Синтез АТФ сопряжён с работой дыхательной цепи
Дыхательная цепь состоит из четырёх комплексов
Комплексы — это гигантские белки. Они пронизывают внутреннюю мембрану митохондрий. Первый и второй комплекс принимают на себя электроны от восстановленных коферментов: НАДH и ФАДH2. При этом на первый комплекс попадают электроны с НАДH, а на второй комплекс с ФАДH2. Дальше мы поговорим об этом подробнее. Пока давайте ответим на простой вопрос: «Как заставить электроны перемещаться по дыхательной цепи?».
Электронам не выгодно прыгать с одного комплекса на другой. Им нет дела до создания градиента концентрации протонов. Митохондрии нужно заставить их перемещаться вдоль этой цепи. Для этого в комплексах дыхательной цепи находятся: коферменты, железо-серные белки, цитохромы и медь. Их строение будет разобрано далее. Но как они заставляют электроны передвигаться? С помощью редокс потенциалов. У них есть и другое название — окислительно-восстановительные потенциалы.
Дело тут такое, у молекул различное сродство к электронам. Чем больше окислительно-восстановительный потенциал молекулы, тем больше у неё сродство к электронам. Если расположить молекулы в порядке возрастания их потенциалов, то мы заставим двигаться электроны вдоль дыхательной цепи. На каждом комплексе у нас будет молекула, которая любит электроны больше, чем на предыдущем. Это заставит их работать! Вот редокс потенциалы для разных молекул.

Мы видим, что максимальный редокс потенциал у кислорода. У остальных молекул поменьше. Они будут располагаться в дыхательной цепи именно в такой последовательности — возрастания окислительно-восстановительных потенциалов.
У разных молекул — разные редокс потенциалы.
Комплекс 1 — НАДH-дегидрогеназа
Этот комплекс состоит из 45 полипептидных цепей! В его состав входит ФМН и железо-серные белки. Его основная задача — принять электроны от НАДH и переместить на убихинон (коэнзим Q10). Комплекс похож на перевёрнутую букву L: часть выходит в матрикс, а другая находится в мембране.
Восстановленный НАДH — из общего катаболизма — отдаёт гидрид ион (два электрона и протон) этому комплексу. Электроны сначала попадают на ФМН, а потом на железо-серные белки — всего их восемь. Я не буду рисовать их все, а нарисую только пару, последний — N-2.
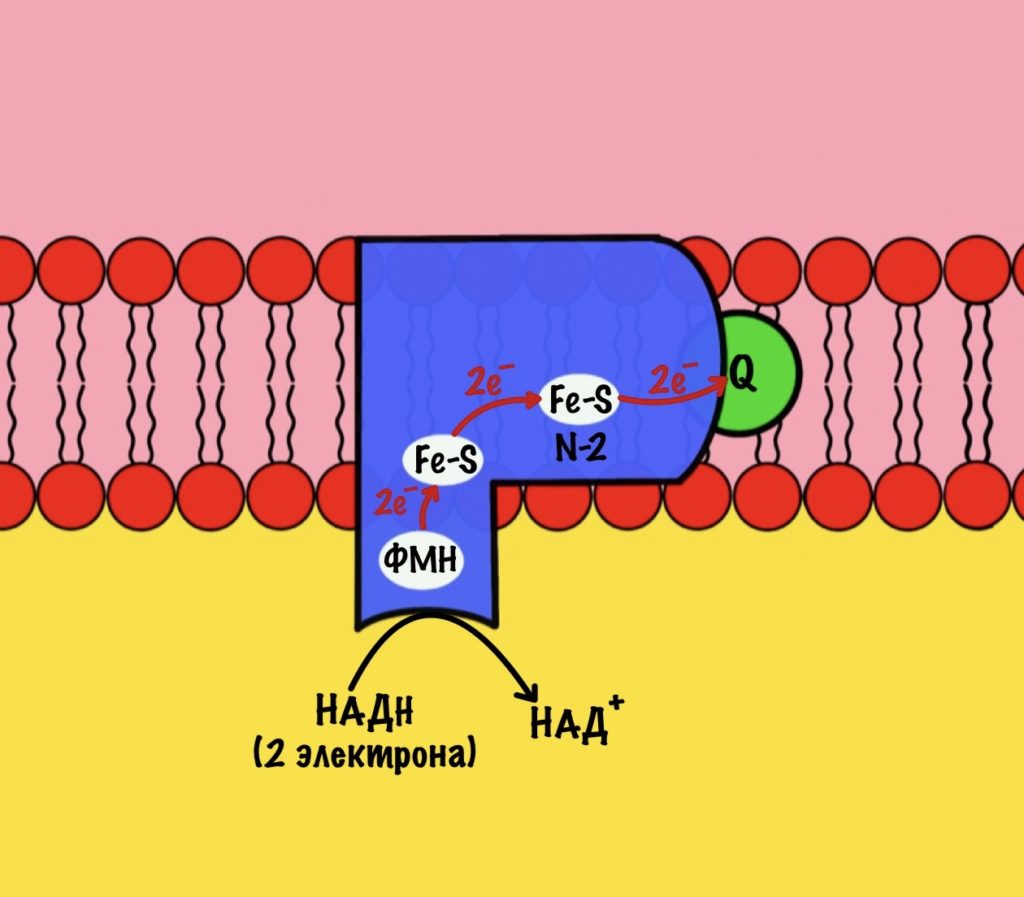
Давайте посмотрим на строение железо-серных белков. Они могут включать один атом железа, два или четыре. Самые простые включают один атом железа — оно связано с четырьмя -SH группами. Железо-серные белки c двумя и четырьмя атомами железа связаны не только с остатками цистеина, но и неорганической серой.
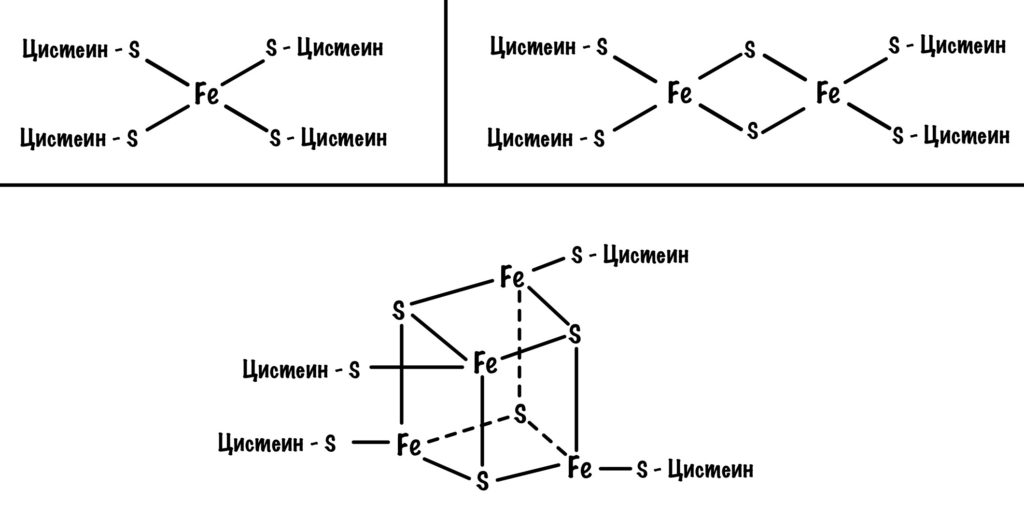
Железо в этих белках имеет заряд +3, но оно может присоединить электрон и перейти в +2. Это и происходит, когда электроны передвигаются от одного железно-серного белка к другому. Чем больше атомов железа в таких белках, тем больше электронов они могут присоединить к себе. Они что-то вроде склада электронов.
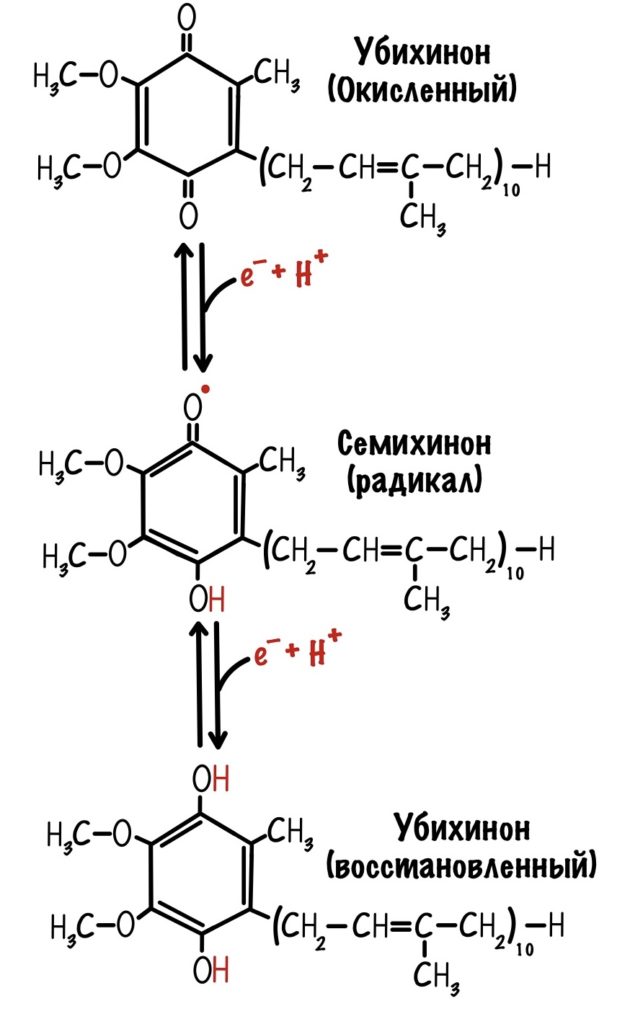
Последний железо-серный белок — N-2. Этот белок отдаёт электроны на убихинон. Убихинон — это жирорастворимая молекула, которая находится во внутренней мембране. Она присоединяется к первому комплексу и ждёт, пока комплекс отдаст ей электроны. Также он присоединяет два протона — один от НАДH, а другой из матрикса митохондрий. Вот как выглядит эта молекула. Убихинон переходит в восстановленную форму.
Мы видим то, о чём говорили в начале — серию окислительно-восстановительных реакций. Электроны из НАДH, через серию переносчиков первого комплекса, попадают на коэнзим Q10. В ходе этой серии реакций выделяется энергия. Энергия идёт на транспорт четырёх протонов из матрикса в межмембранное пространство. Точный механизм транспорта неизвестен, но он подтверждён практически. Так что первый комплекс — протонная помпа.
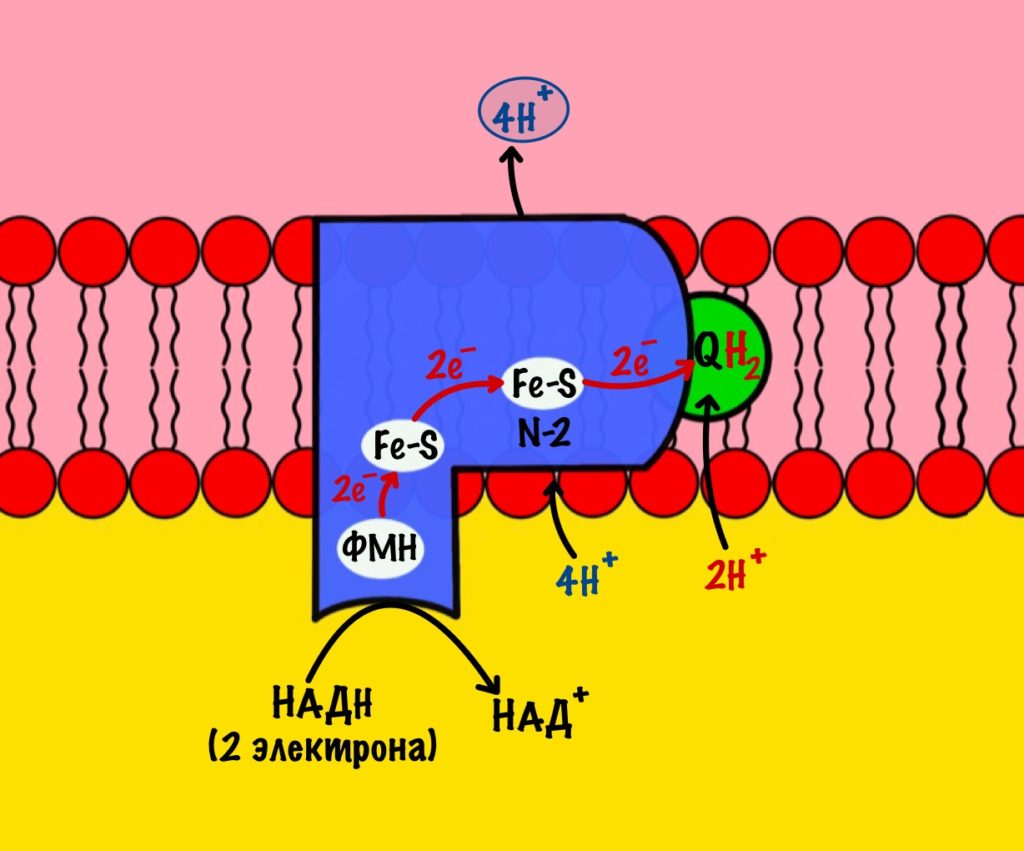
Первый комплекс переносит два электрона с НАДH на коэнзим Q10. Энергия окислительно-восстановительных реакций используется на транспорт четырёх протонов из матрикса в межмембранное пространство.
Далее убихинон отсоединяется от первого комплекса и путешествует по внутренней мембране. Он жирорастворимый, поэтому это не составляет ему труда. Восстановленный убихинон присоединится к третьему комплексу, которому и отдаст свои электроны и протоны.
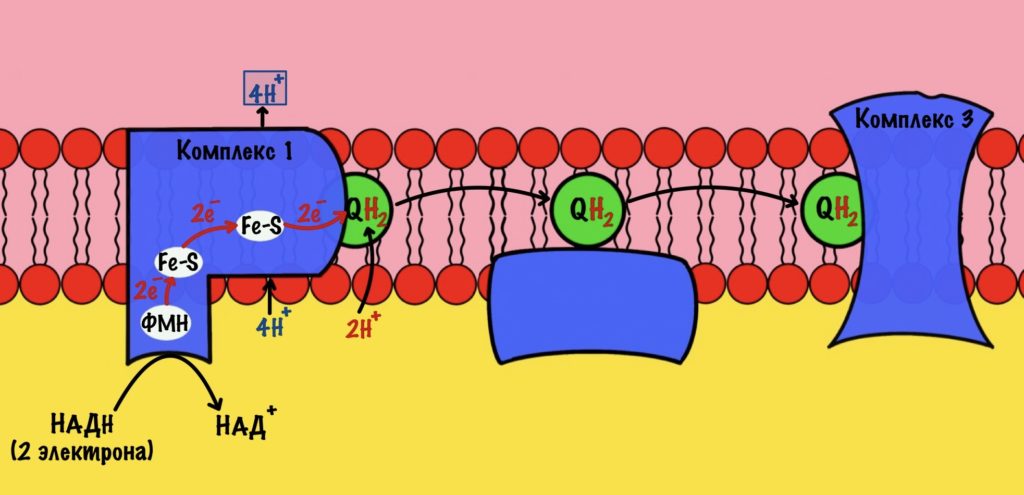
Стоп, а где второй комплекс? Их же всего четыре, почему убихинон переносит электроны сразу на третий? А сейчас разберёмся.
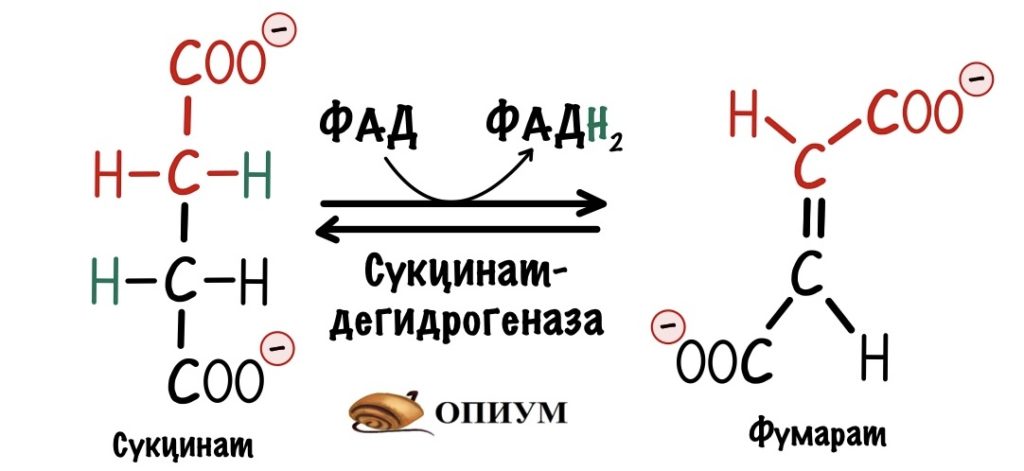
Второй комплекс — сукцинатдегидрогеназа
Мы уже встречались с ним — в реакциях цикла Кребса! Сукцинатдегидрогеназа катализировала превращение сукцината в фумарат. Кофермент этой реакции — ФАД. Это единственный фермент из цикла трикарбоновых кислот, который крепится к внутренней мембране митохондрий.
Задача второго комплекса — транспорт электронов с ФАДH2 их на убихинон. Второй комплекс намного меньше первого, он включает четыре субъединицы. Только одна ответственна за реакцию сукцинатдегидрогеназы — субъединица А. Кофермент ФАД тесно связан с этим комплексом. Остальные субъединицы транспортируют электроны. В комплексе есть три железо-серных белка и цитохром b. Железо-серные белки переносят электроны на убихинон. Цитохром b не участвует в транспорте электронов, он нужен для избежания утечек электронов. Иногда электроны сбиваются со своего пути, а это может привести к образованию токсичных форм кислорода. Это радикалы, которые могут натворить много неприятных дел внутри клетки. Так что цитохром b на подсейве.
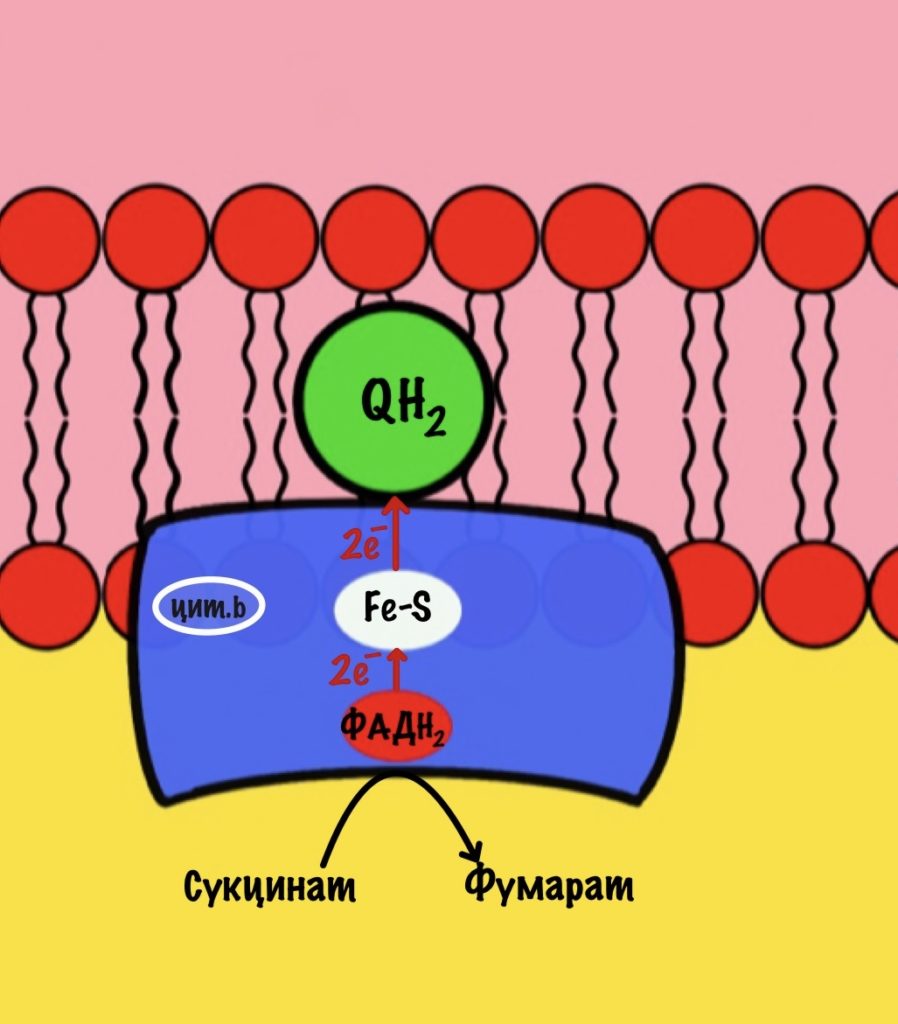
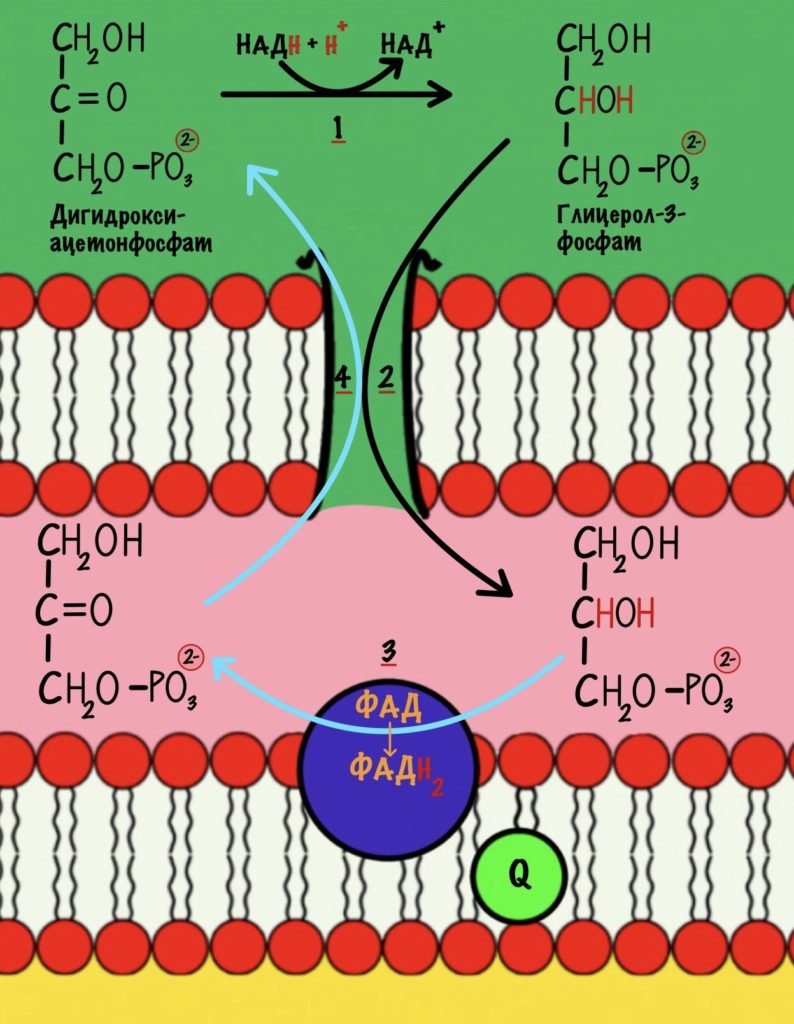
Убихинон принимает электроны с ФАДH2 не только от сукцинатдегидрогеназной реакции. Например, он может присоединять электроны, которые образовались в глицеролфосфатном челноке.
Второй комплекс не работает, как протонная помпа. Он не транспортирует протоны из матрикса в межмемрбанное пространство.
Второй комплекс переносит два электрона от ФАДH2 на убихинон. Энергия окислительно-восстановительных реакций не используется на транспорт протонов.
Дальше убихинон будет путешествовать по мембране, пока не присоединится к третьему комплексу. Там он отдаст электроны и протоны.
Третий комплекс — цитохром bc1 комплекс
Мы ещё не встречались с цитохромами, что это такое? Цитохромы — это белки, которые содержат в себе гем. Гем состоит из порфирина, который соединен с железом. Порфирин состоит из четырёх пятичленных колец. В каждом кольце есть атом азота, с помощью которого кольца соединяются с атомом железа. У порфирина могут быть разные радикалы, по ним цитохромы делятся на разные типы.
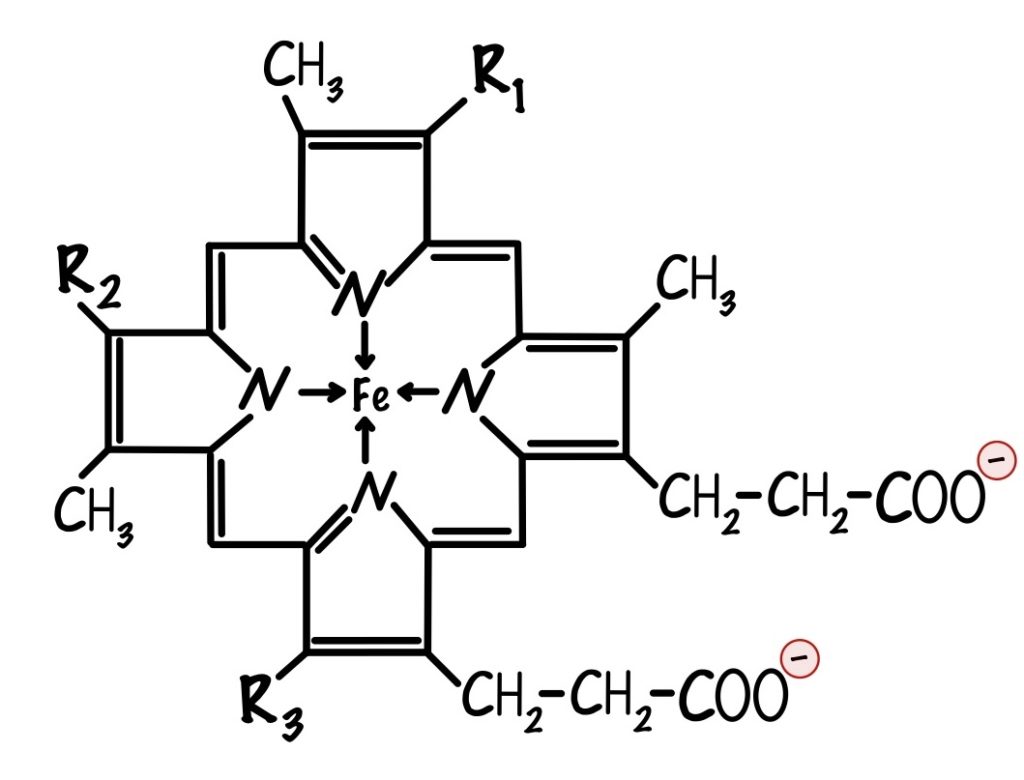
Важно понять, что в цитохромах есть один атом железа. Он может принимать электрон и менять степень окисления: из +3 в +2. Цитохромы могут принимать только один электрон, потому что в них один атом железа.
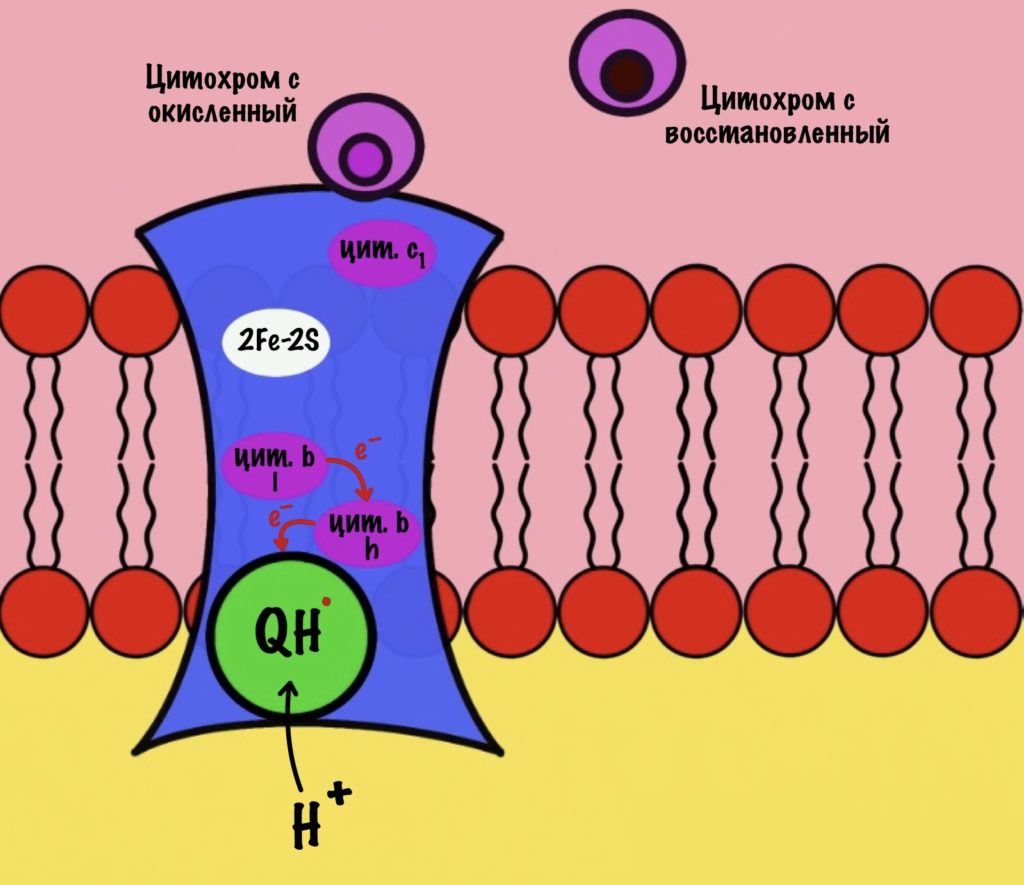
Третий комплекс состоит из двух субъединиц. В каждой субъединице есть: 2Fe-2S железо-серный белок, цитохром b, цитохром c1. В цитохроме b есть два порфириновых кольца, поэтому его делят на l и h. К субъединицам крепится цитохром c — он находится в межмембранном пространстве митохондрии. Цитохром c растворим в воде, в отличие от остальных цитохромов.
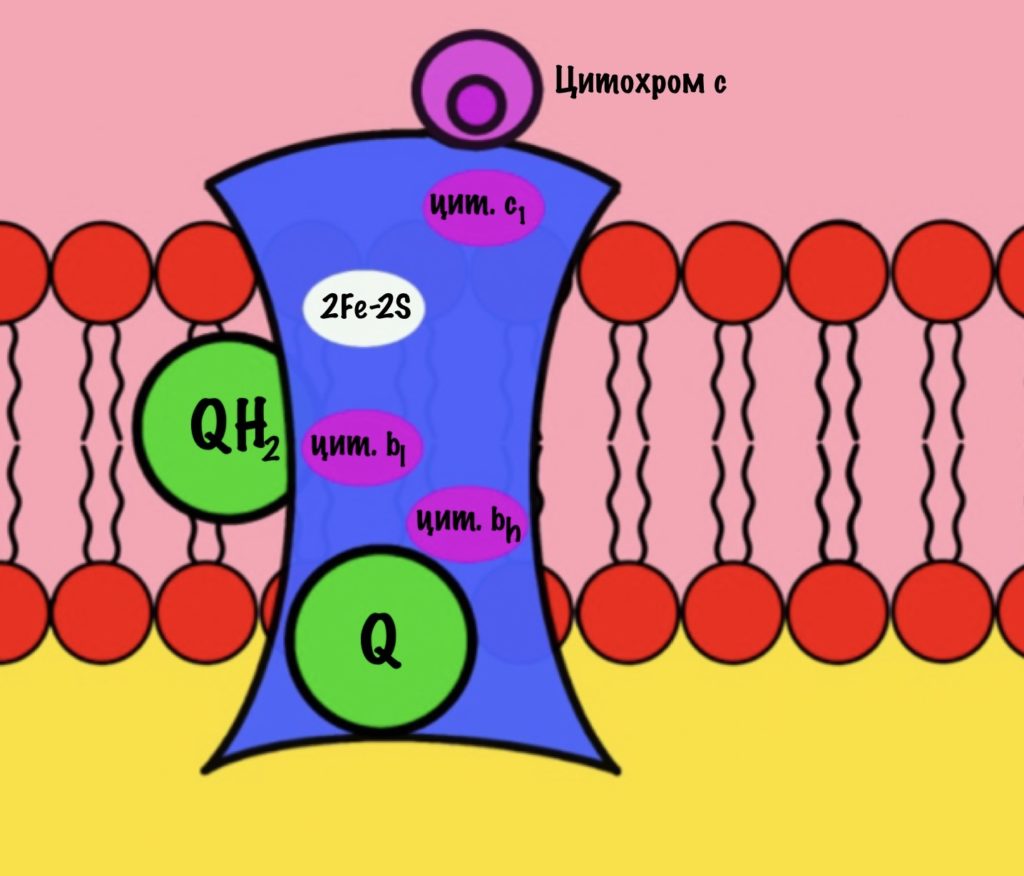
Задача этого комплекса — транспорт электронов с восстановленного убихинона на цитохром c. Разберём на примере одной субъединицы.
К комплексу присоединяется восстановленный убихинон. Он отдаёт один электрон на цитохром b, а другой на железо-серный белок. Дальше железо-серный белок отдаст электрон цитохрому c1. Почему только один? Вспоминаем, что железо может присоединить только один электрон — перейти в +2. Поэтому нужна такая развилка для электронов. В это же время от убихинона отщепляются два водорода, которые транспортируются в межмембранное пространство третьим комплексом. Убихинон становится окисленным и отсоединяется от третьего комплекса.
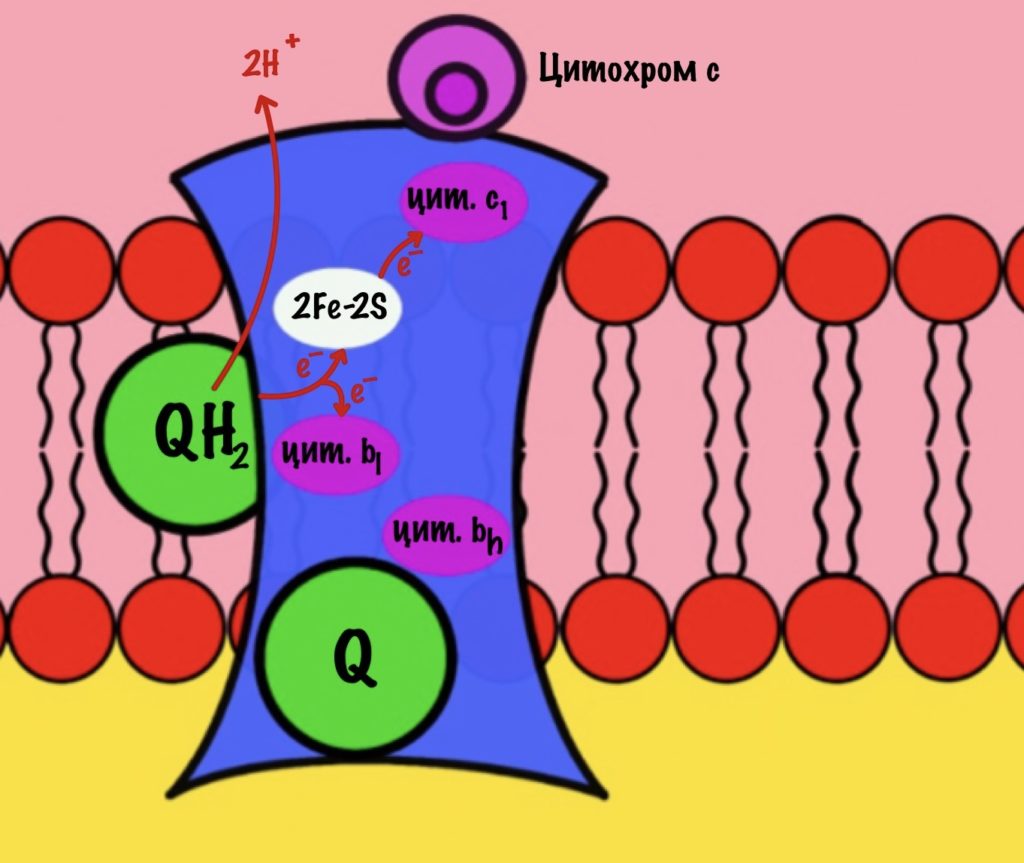
Цитохром c1 передаёт электрон цитохрому c. Тот говорит: «до связи». И отчаливает от третьего комплекса. Дальше восстановленный цитохром c попадёт на четвёртый комплекс, но об этом пока рано. К третьему комплексу присоединится следующий окисленный цитохром c.
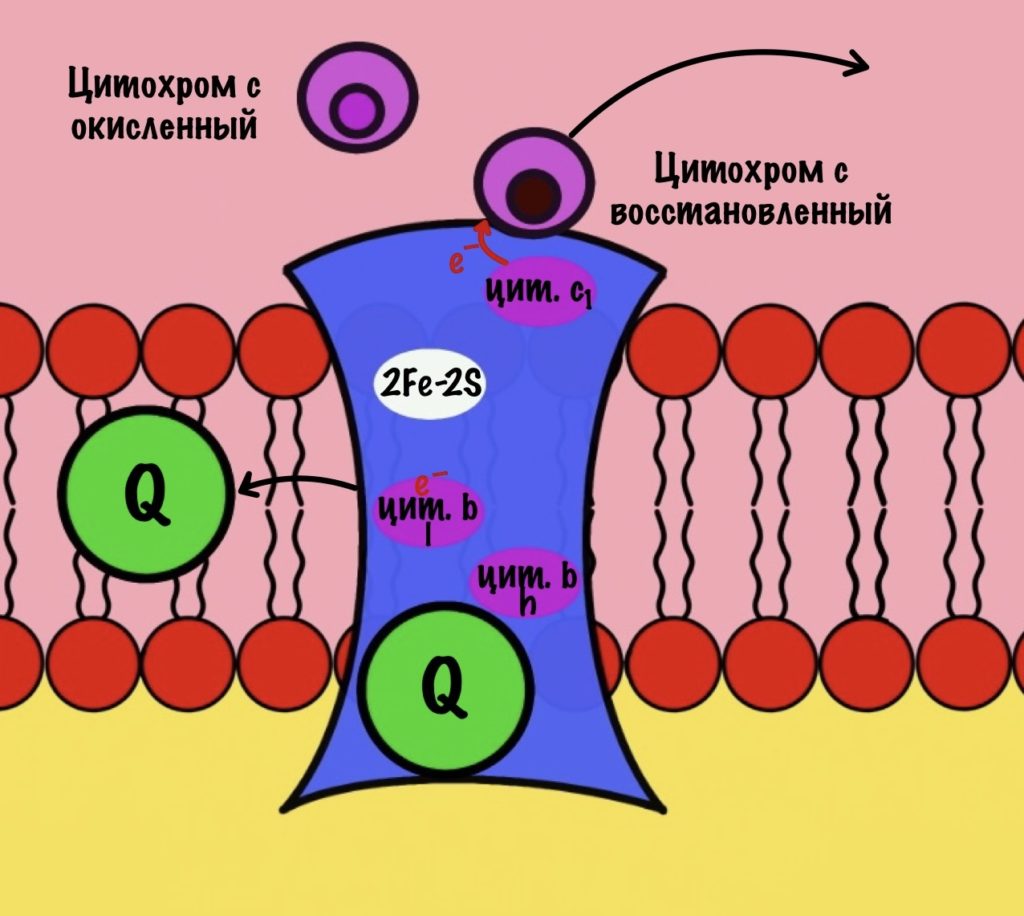
Что будет со вторым электроном? У цитохрома b есть два порфирина с железом: l и h. Электрон переходит с порфирина l на h, а дальше падает на убихинон. Но не тот же самый, а другой — окисленный. Он сидел в третьем комплексе и ждал своего часа. Также убихинон присоединяет протон водорода из матрикса — образуется полувосстановленный убихинон (семихинон). С этой молекулой пока ничего не сделаешь.
К третьему комплексу присоединяется ещё один восстановленный убихинон. Он снова отдаёт один электрон железо-серному белку, а другой цитохрому b. Два протона от убихинона вылетают в межмембранное пространство. От железо-серного белка электрон попадает на цитохром c1, а потом и на цитохром c. Ещё один электрон отправился на четвёртый комплекс.
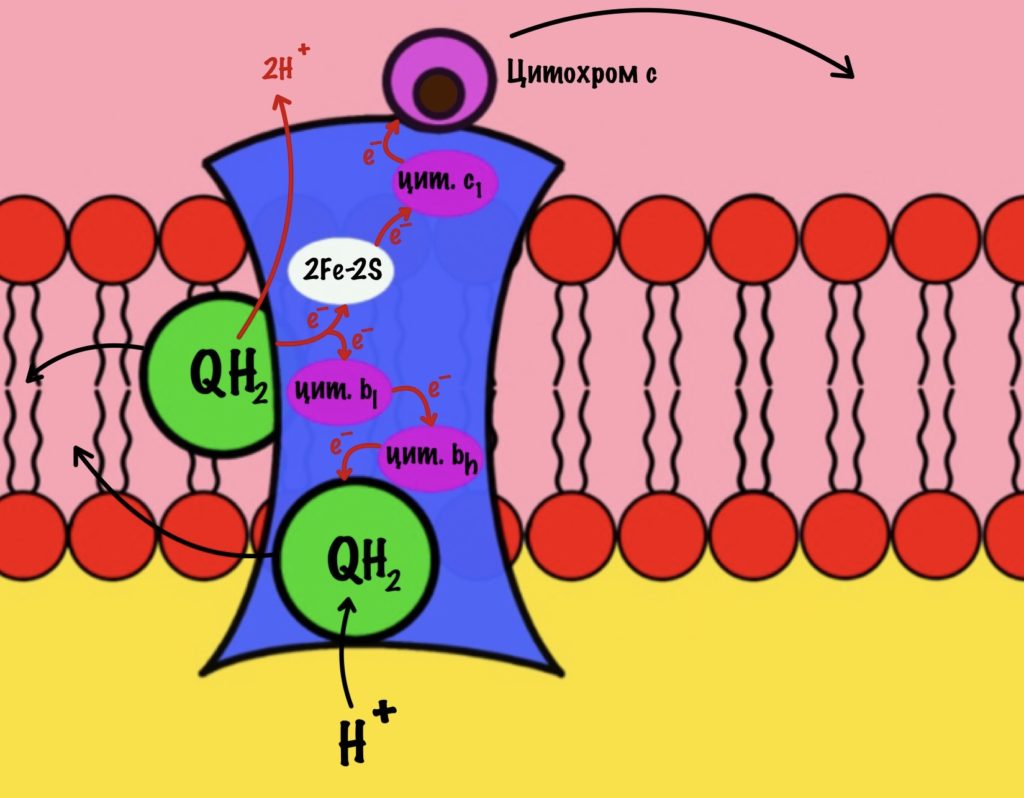
Второй электрон и протон из матрикса присоединяются к полувосстановленному убихинону, который ждёт своего часа. Он переходит в восстановленный убихинон. Восстановленный убихинон отсоединяется от третьего комплекса, чтобы присоединиться в другом месте к третьему комплексу))) Цикл завершён, у него даже есть своё название — цикл Q или убихиноновый цикл.
Третий комплекс переносит два электрона от убихинона на два цитохрома c. Энергия окислительно-восстановительных реакций используется на транспорт четырёх протонов в межмембранное пространство.
Четвёртый комплекс — цитохром c оксидаза
Цитохром с оксидаза имеет димерную структуру. Каждый мономер состоит из 13 субъединиц. Его задача — перенос электронов от цитохрома c на кислород. Кислород — это финальный акцептор электронов в дыхательной цепи.
Второй мономер отвечает за связывание цитохрома c и присоединение его электрона. Электрон принимают на себя два атома меди, которые связаны ковалентной связью друг с другом. К четвёртому комплексу атомы меди присоединяются с помощью цистеина, гистидина и метионина — это напоминает железо-серные белки. За счёт такого строения эти два атома могут принять только один электрон. Я запишу эти ионы вот так — CuA/CuA.
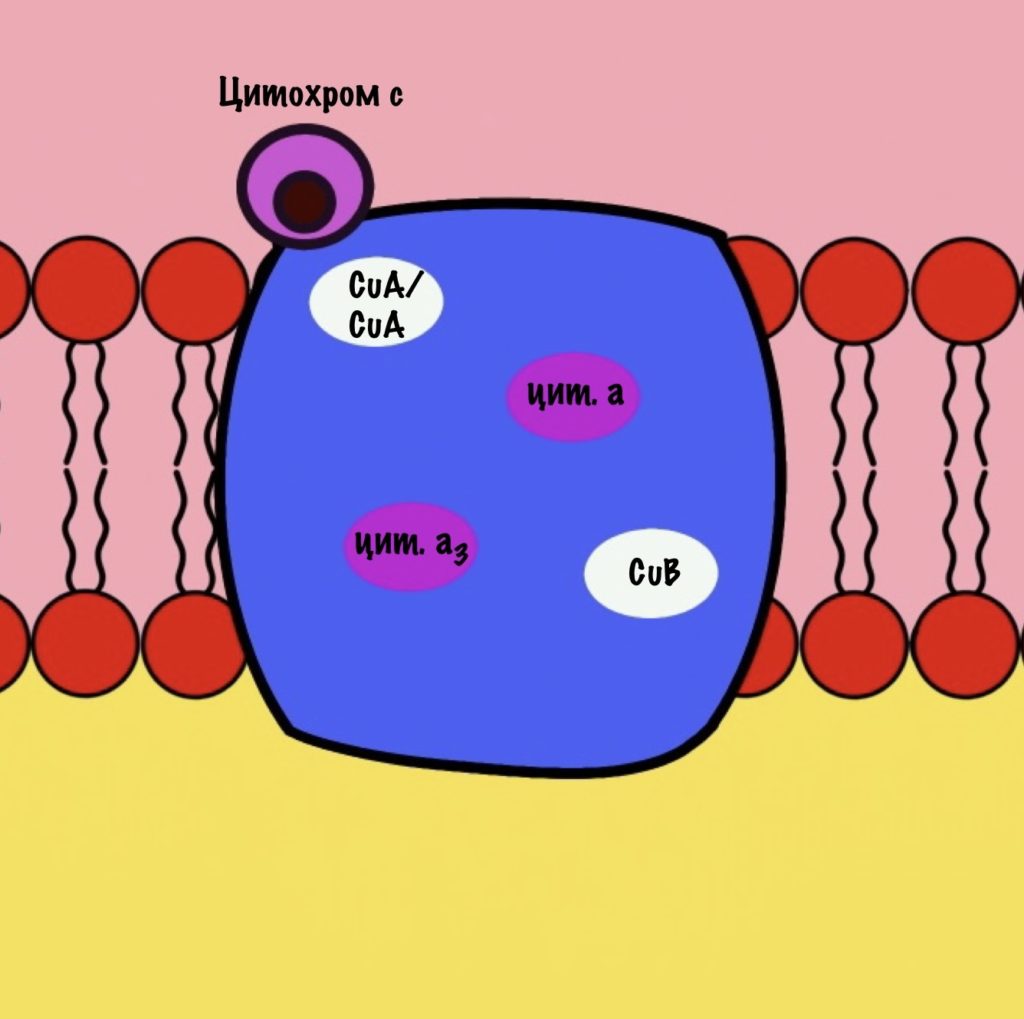
Первый мономер отвечает за передачу электронов на кислород. Он состоит из двух цитохромов: a и a3. Ещё в нём есть один ион меди +2 — CuB. Медь связана с цитохромом a3 (хоть на рисунке это и не видно).
Цитохром c присоединяется к четвёртому комплексу и передаёт свой электрон на CuA/CuA. Далее атомы меди отдают электрон на цитохром a, а тот на цитохром a3. Они могут принять только по одному электрону — там одно железо +3. Цитохром a3 скидывает электрон на CuB. Медь меняет степень окисления — переходит из +2 в +1.
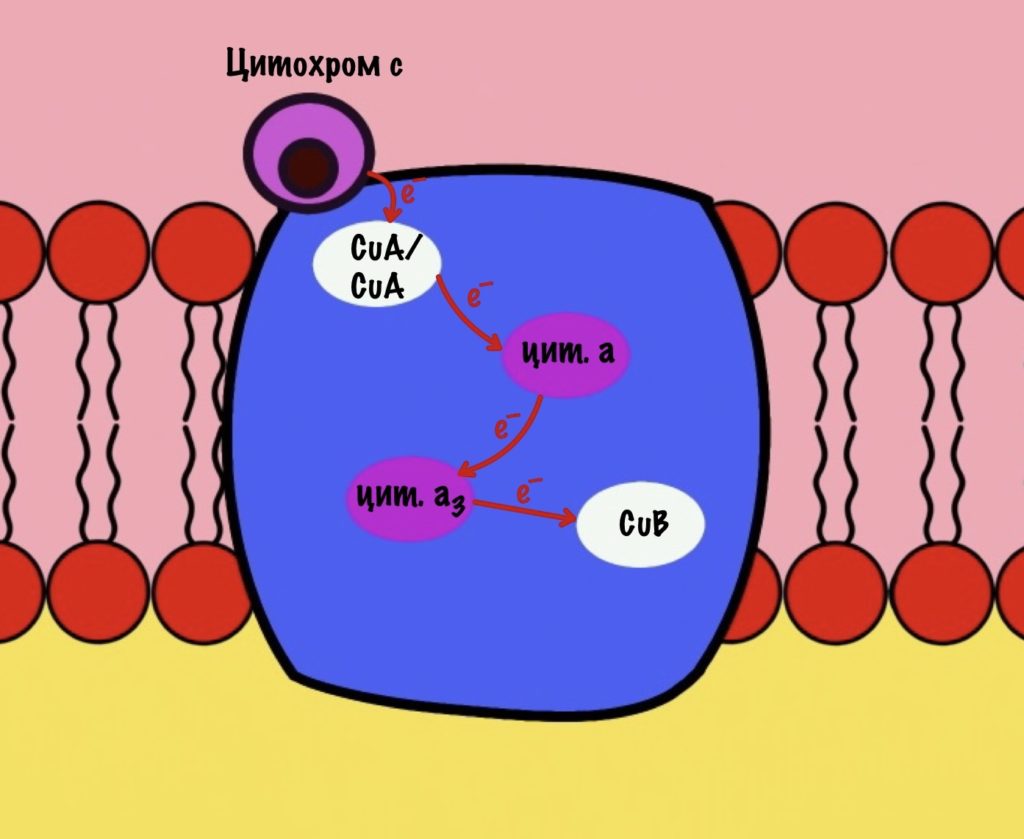
Дальше присоединяется второй цитохром c. Повторяется всё, что я написал выше. Но электрон не может попасть на CuB, потому что она уже +1, а больше электронов принять не может. Поэтому электрон останавливается на цитохроме a3. Теперь цитохром a3 и ион меди +1 могут связаться с кислородом — формируется пероксидный мостик. Кислород поглощает два электрона: один с цитохрома a3, а другой с CuB.
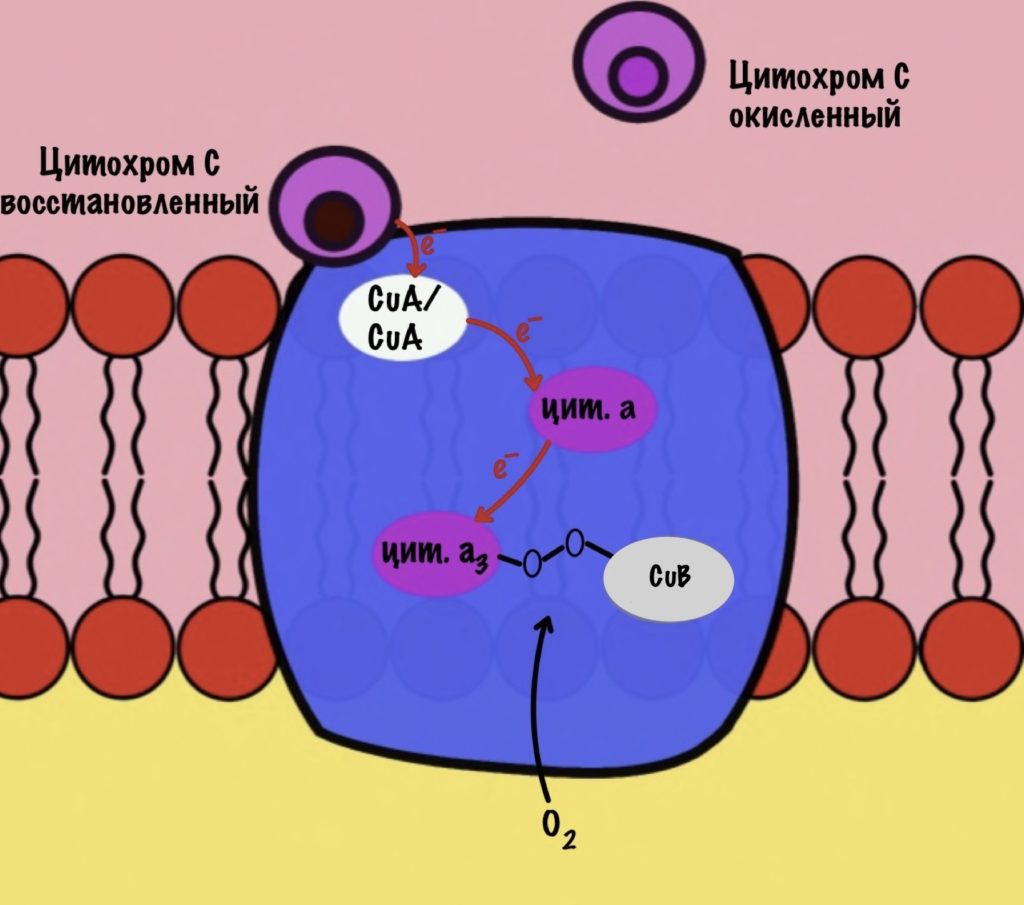
Присоединяется третий цитохром c. У меня ощущение, что я рассказываю анекдот с затянутой концовкой. Этот электрон используется для того, чтобы разрушить пероксидный мостик между железом и медью. Но нужен не только электрон, но и протон — он берется из матрикса митохондрии. Это приводит к разрушению пероксидного мостика. Остаётся цитохром а3, который связан с кислородом, и медь с гидроксильной группой.
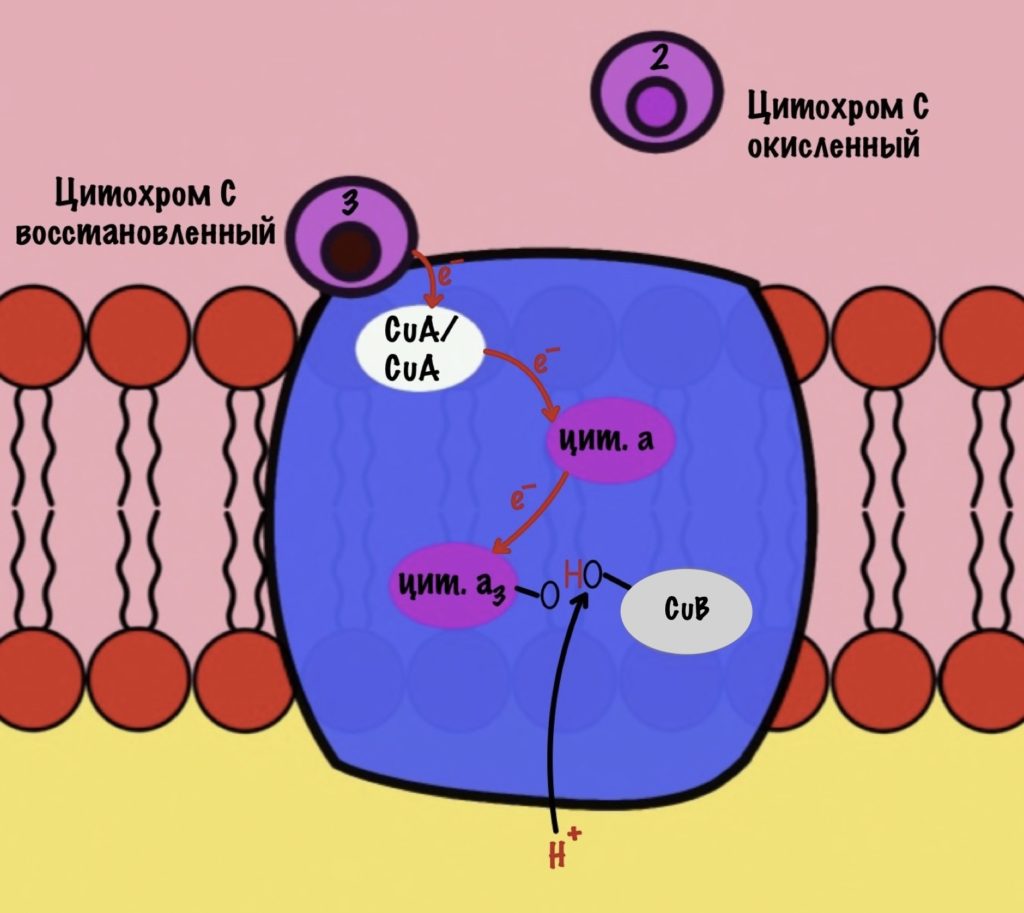
Присоединяется четвёртый электрон от цитохрома с, а один протон берется из матрикса. Это приводит к образованию гидроксильной группы на цитохроме а3.
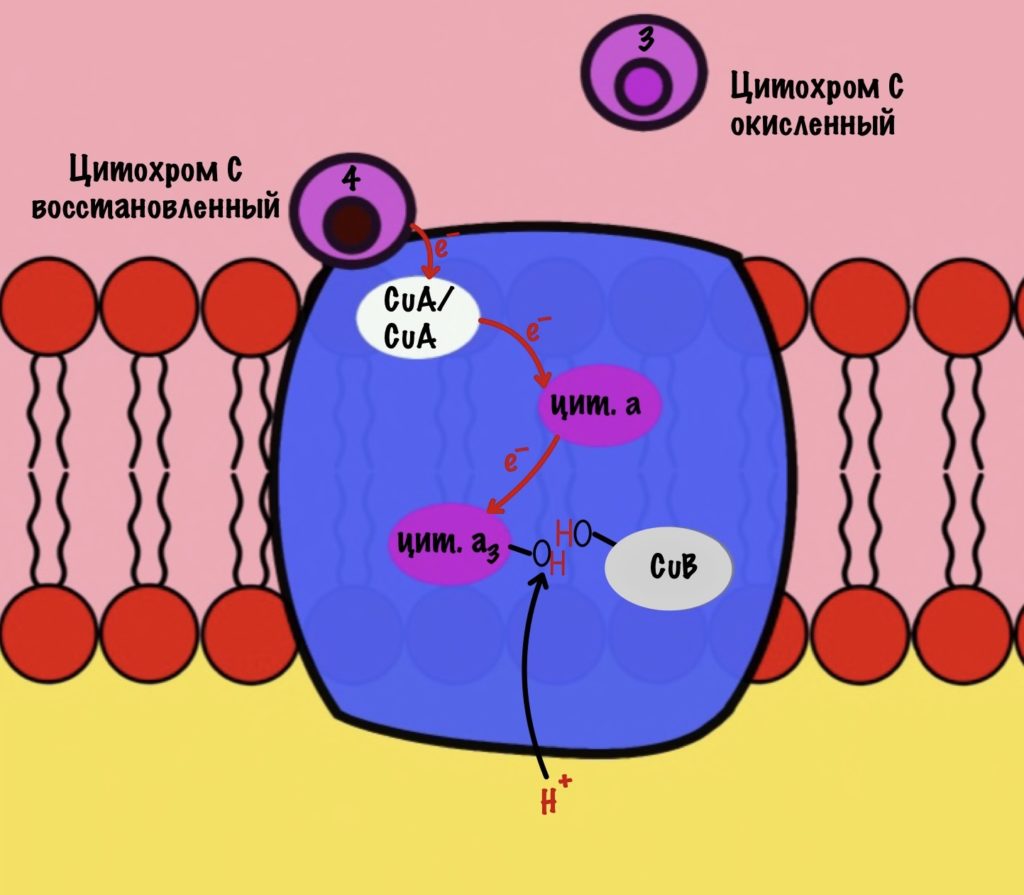
Из матрикса берутся ещё два протона — это приводит к образованию двух молекул воды. При этом цитохром а3 и медь возвращаются в свою начальную форму.
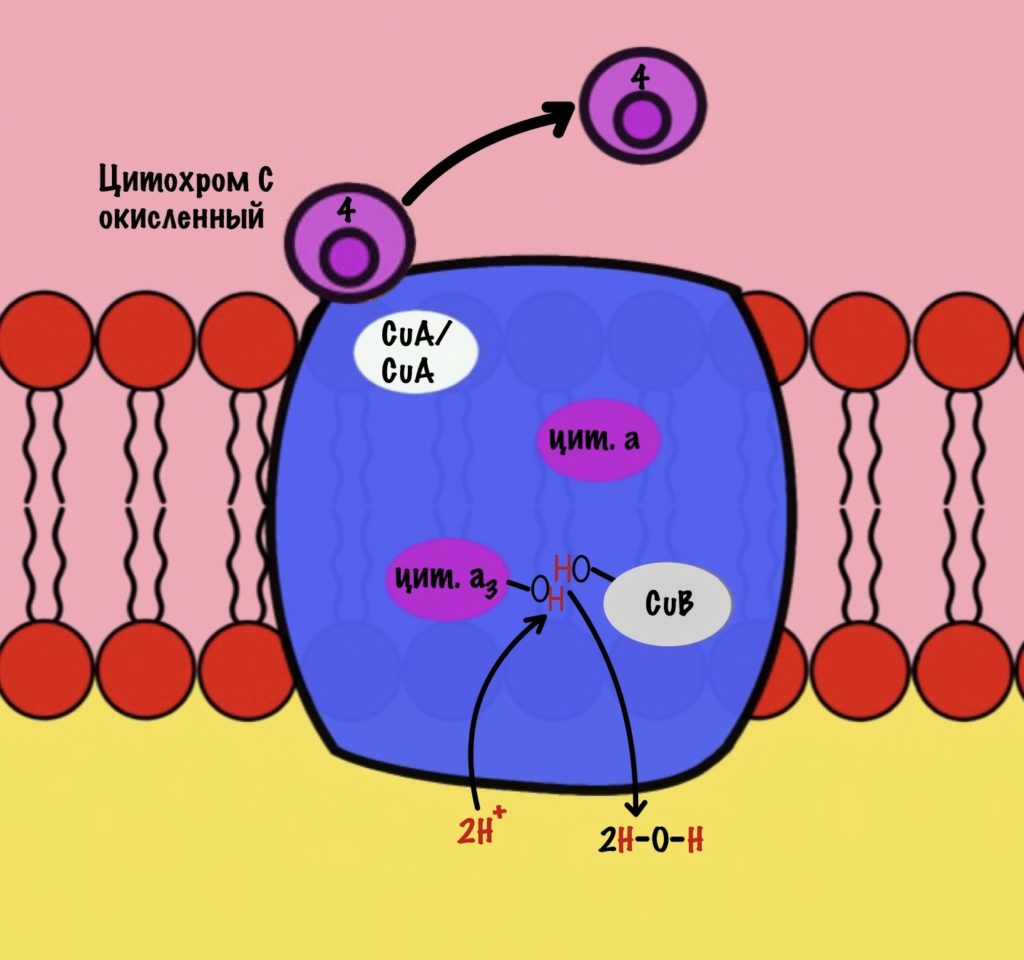
Обсудим, что у нас вышло. Мы присоединили четыре электрона к четвёртому комплексу, а также взяли из матрикса четыре протона водорода и кислород. В итоге мы получили две молекулы воды, да ещё избавились от опасных электронов. Они могут много чего натворить в клетке. Энергия окислительно-восстановительных реакций одного электрона используется на транспорт одного протона из матрикса в межмембранное пространство. Всего использовали четыре электрона — четыре протона вылетело в межмембранное пространство.
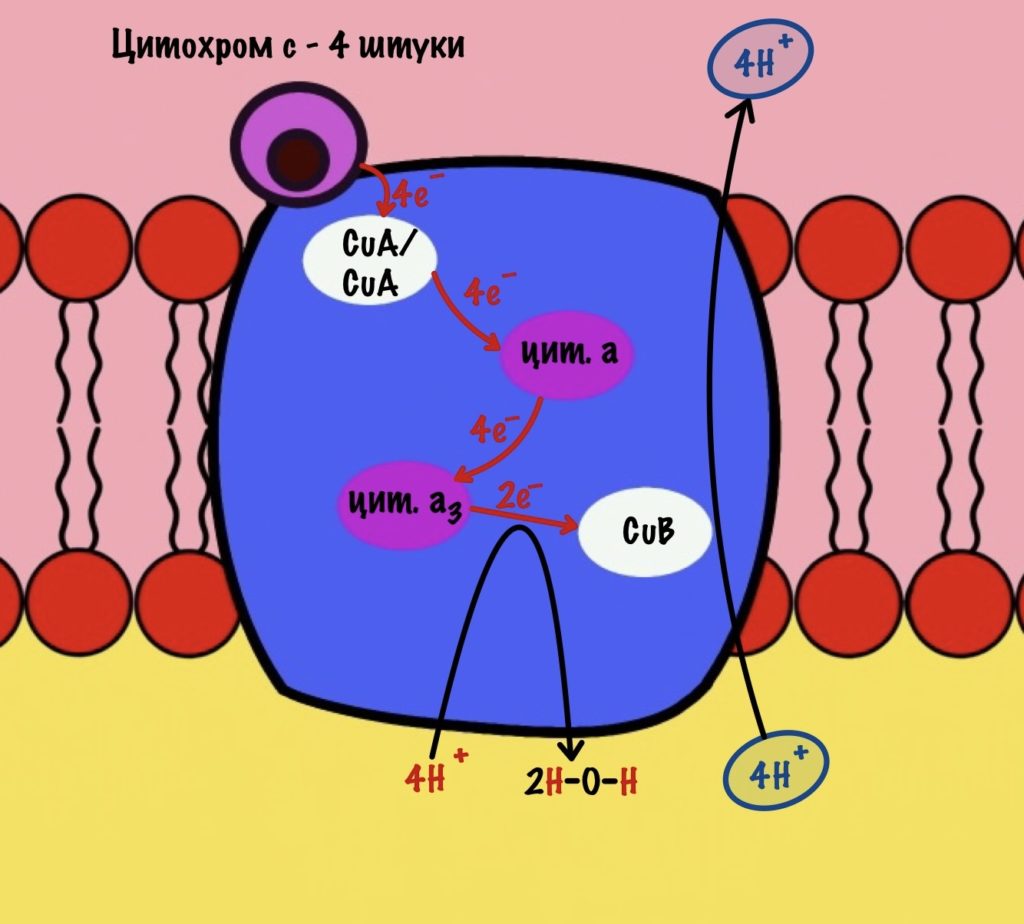
Воспоминаем, что до этого мы использовали только по два электрона во всех комплексах. Да и НАДH и ФАДH2 несли себе всего по два электрона. Не проблема, просто поделим всё на два. И у нас получится:
Четвёртый комплекс переносит два электрона на 1/2 кислорода — образуется молекула воды. Энергия восстановительных реакций используется для транспорта двух протонов водорода.
Общий подсчёт: сколько протонов транспортируется из матрикса за счёт двух электронов?
В ходе работы первого комплекса были транспортированы 4 протона. Третьего — 4 протона. А четвёртого — 2 протона. Второй комплекс протоны не транспортирует. Первый комплекс присоединяет НАДH, а второй — ФАДH2. Второй комплекс передаёт электроны на третий комплекс, пропуская первый. Получается, что энергия электронов одной молекулы НАДH используется на транспорт 10 протонов. А энергия электронов одной молекулы ФАДH2 на транспорт 6 протонов.
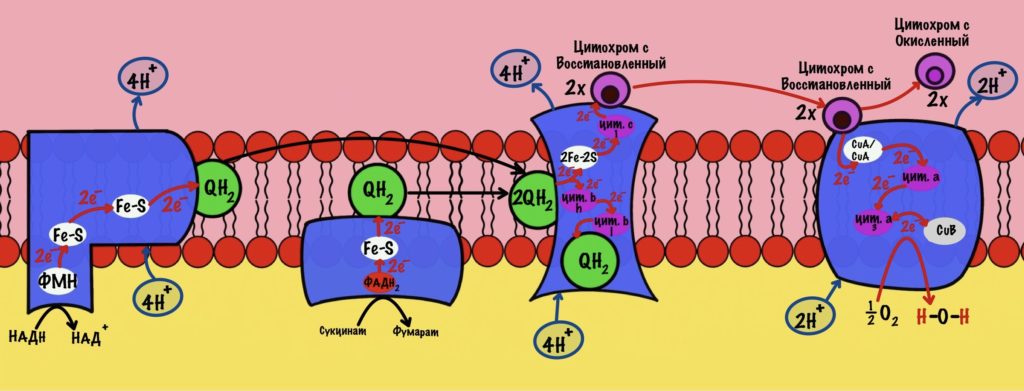
Энергия электронов 1 НАДH идёт на транспорт 10 протонов.
Энергия электронов 1 ФАДH2 идёт на транспорт 6 протонов.
Теперь нам нужно превратить эти протоны в энергию.
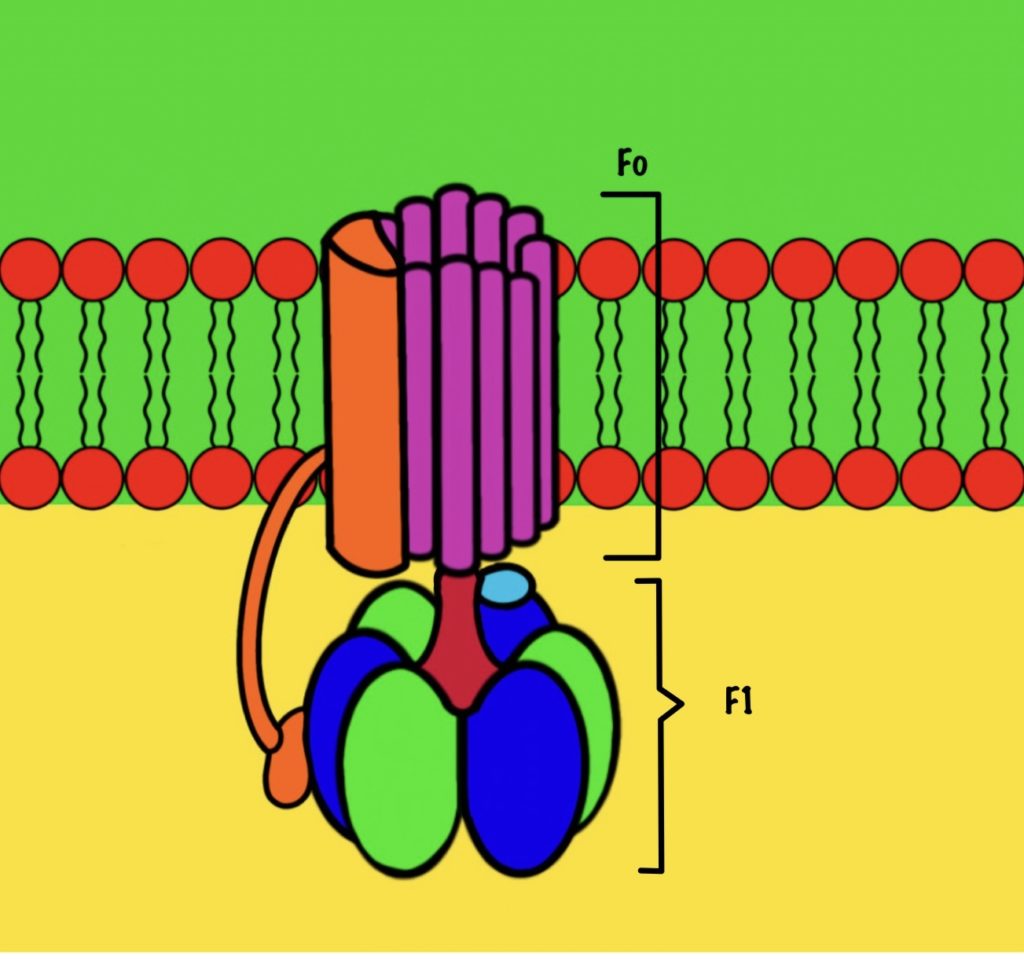
АТФ-синтаза
АТФ-синтаза — это фермент, который состоит из двух больших частей (или доменов): Fо и F1. Fо — это протонная помпа, которая встроена в мембрану. Если перевести название на русский — олигомицин-чувствительная фракция (часть). F1 — это каталитический домен, который синтезирует АТФ. F1 — первая фракция (часть).
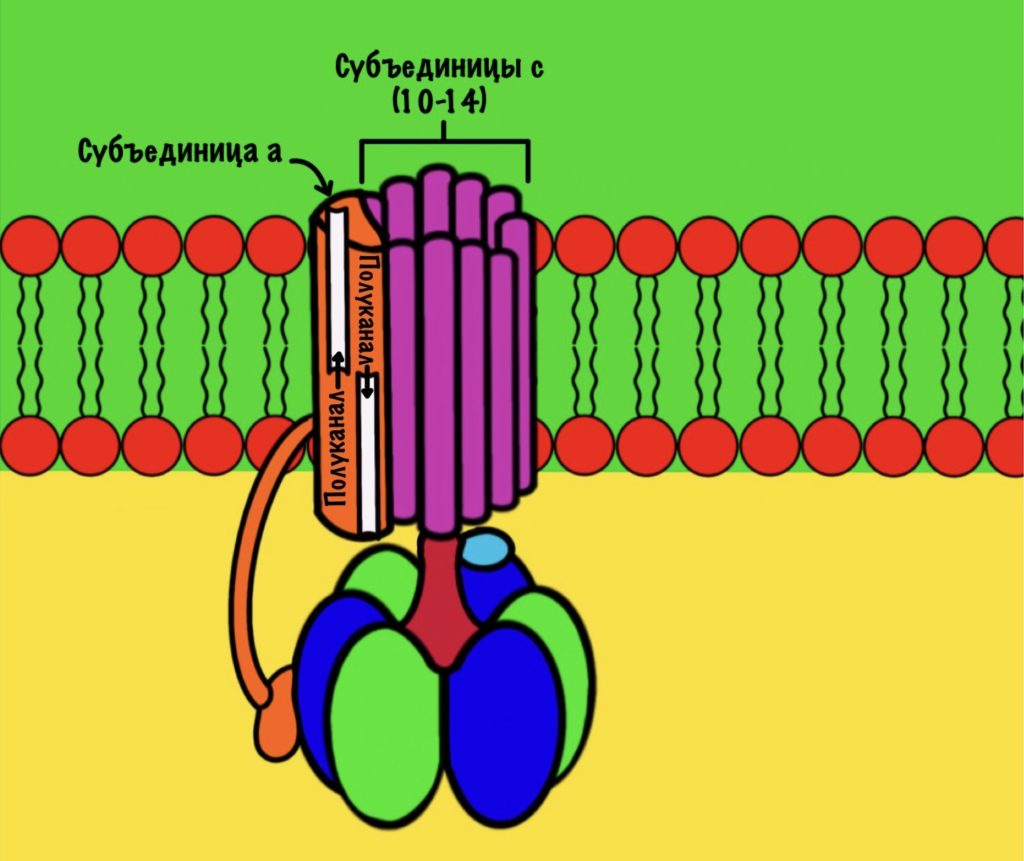
F0 домен состоит из 10-14 субъединиц c и одной субъединицы a. C субъединицы образуют жёсткое кольцо — оно вращается, когда присоединяет протон водорода. В субъединице a есть полуканалы для протонов. Именно полуканалы, а не целые. Один полуканал связан с межмембранным пространством, а второй с матриксом. Они отделены друг от друга.
F1 домен состоит из пяти типов полипептидных цепей — альфа, бета, гамма, эпсилон и дельта. Три альфы и три беты образуют гексамер — похож на клевер. Он ответственен за синтез АТФ. Этот гексамер связан с 10-14 субъединицами с с помощью гамма и эпсилон цепей. Две эти цепи образуют ножку, которая вращается вместе с субъединицами c. Наибольшая роль здесь у гамма субъединицы. Вы можете подумать, что эта ножка будет вращать гексамер, но это не так! Гексамер всегда остаётся на месте, а вертится только ножка и субъединицы c.
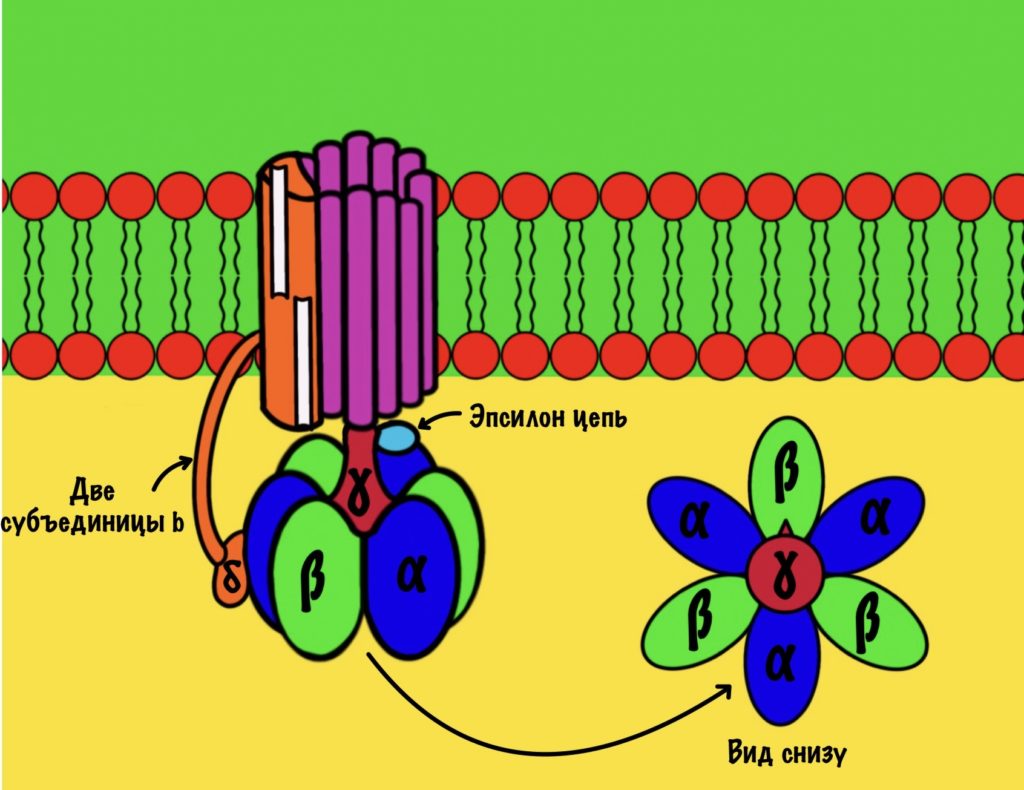
Дельта субъединица стабилизирует гексамер, она не даёт ему развалиться и вращаться вместе с ножкой. Ещё есть две субъединицы b, которые связывают между собой субъединицу a и субъединицу дельта.
Fо и F1 регион связаны через:
- Гамма ножку и эпсилон полипептидную цепь;
- Субъединицы b, которые связывают субъединицы a и дельта.
АТФ синтаза состоит из двух регионов:
- Вращательного — кольцо из субъединиц c и ножки гамма-эпсилон;
- Стационарного: гексамер из 3-альфа и 3-бета цепей, субъединицы a и b, а также дельта-субъединицы.
Регион F1
Начнём с него, а потом перейдём к вращению нашего кольца. Нас интересует гексамер, который участвует в синтезе АТФ. Гексамер состоит из трёх альфа и трёх бета цепей.

При этом альфа цепи всегда связаны с АТФ и не участвуют в его синтезе, поэтому они нас не интересуют. А вот бета цепи ответственны за:
- Связывание АДФ и неорганического фосфата;
- Синтез АТФ;
- Высвобождение АТФ.
Только один из этих процессов связан с вращением c-кольца, угадаете какой? Нет — это не синтез АТФ. Удивительно, но каждая из бета-цепей легко синтезирует АТФ. Они настолько сближают АДФ с фосфатом, а также создают гидрофобное окружение вокруг них, что реакция синтеза АТФ идёт без каких-либо проблем и затрат энергии. А вот высвободить образованную АТФ эти субъединицы не могут, они имеют слишком большое сродство к ней. Поэтому, если мы сравним обычную ферментативную реакцию с получением АТФ, то увидим такое…
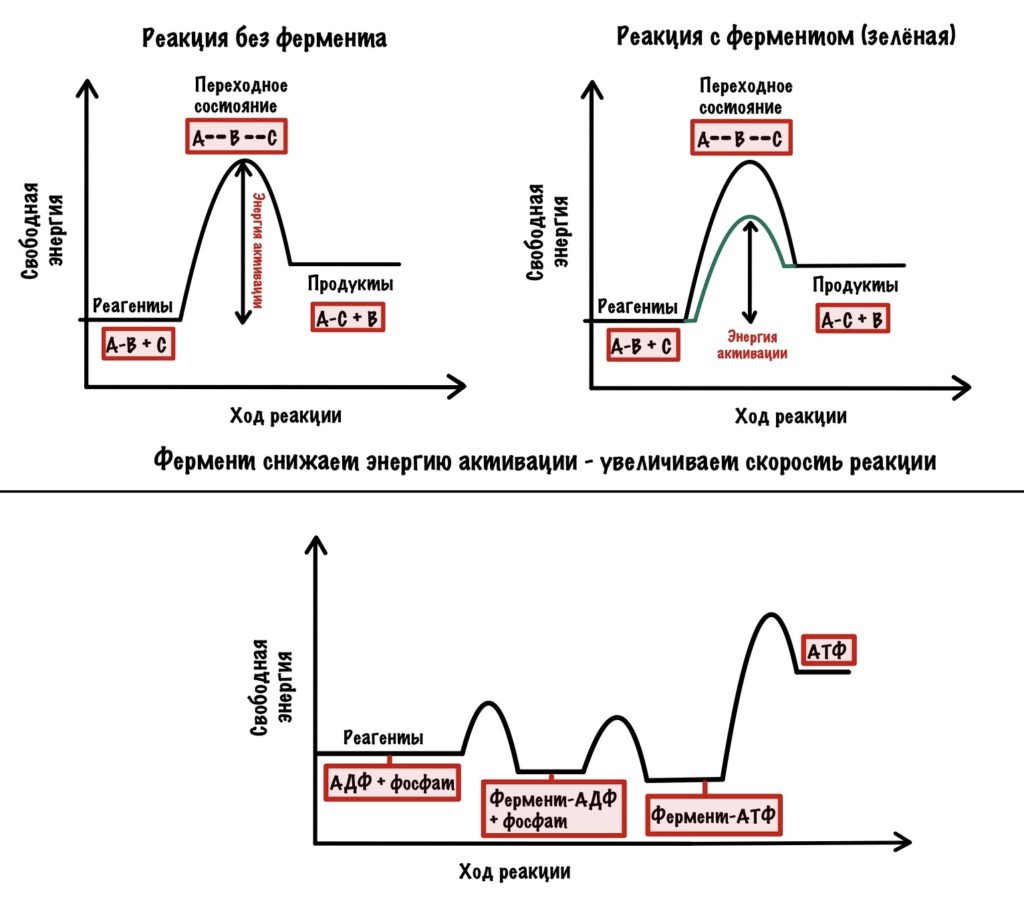
Видите? В образовании АТФ из АДФ и фосфата почти нет изменения свободной энергии, в отличие от ферментативной реакции. Но сколько нужно энергии для её высвобождения — жуть! Для того, чтобы высвободить АТФ нужно вращение, создаваемое c-кольцом. Вращение предоставляет энергию для высвобождения АТФ. Оно вызывает конформационные изменения в бета-субъединицах, благодаря которому они выпускают АТФ из своих лап.
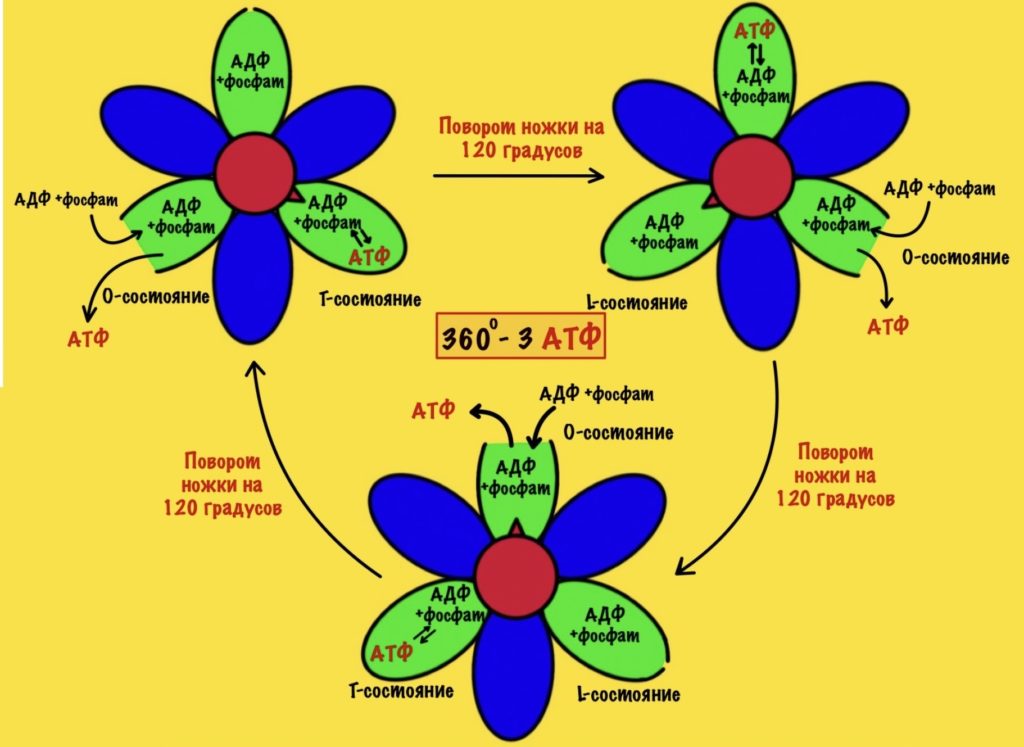
Вернёмся к бета-цепям. Они могут быть в трёх состояниях: open, loose и tense (открытое, свободное и напряжённое). Или O, L и T состояния. В O-состоянии бета-субъединица присоединяет АДФ и неорганический фосфат. В L-состоянии бета закрывает фосфат и АДФ в ловушку — они не могут выбраться из субъединицы. В T-состоянии синтезируется АТФ, которая не может отсоединиться от фермента из-за его высокого сродства.
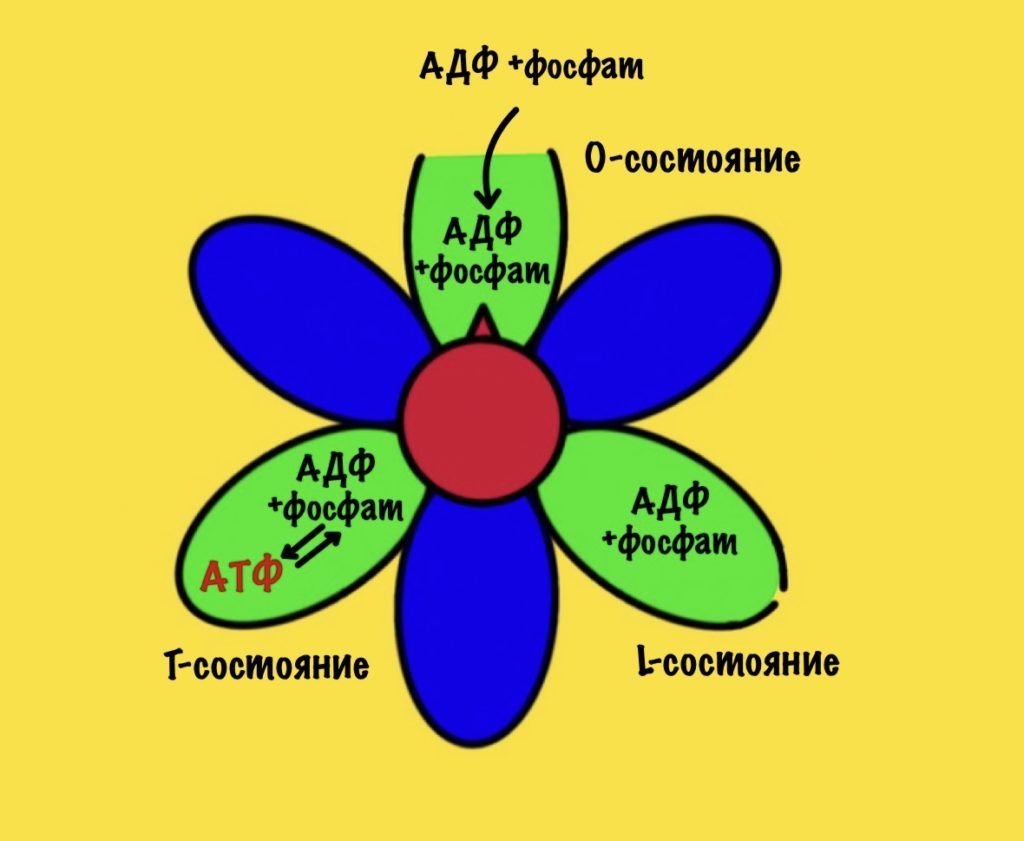
В АТФ-синтазе каждая из бета-субъединиц находится в одном из этих состояний. Одна в O, другая в L, а третья в T. Вращение гамма ножки вызывает переход бета-субъединицы из одного состояния в другое. Получается, что любые две бета-субъединицы не могут существовать в одном состоянии одновременно.
Ножка вращается на 120 градусов по часовой стрелке (если смотреть снизу). Это вызывает конформационные изменения в субъединицах, что приводит к переходу из одного состояния в другое. При этом одна бета-субъединица переходит из O-состояния в L. Вторая из L-состояния в T. А третья из T-состояния в O. В О состоянии бета-субъединица отсоединяет АТФ, а потом связывается со следующим АДФ и неорганическим фосфатом.
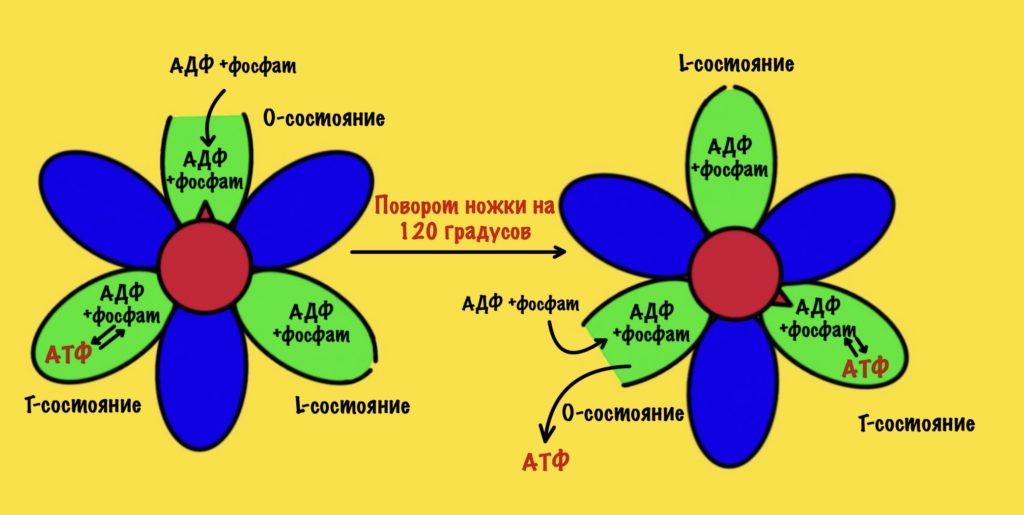
Давайте сделаем полный круг вращения. 360 градусов — это три поворота для нашей ножки. Каждая субъединица изменит свое состояние три раза. Получается, что в ходе такого вращения будет синтезировано три молекулы АТФ.
Регион Fo
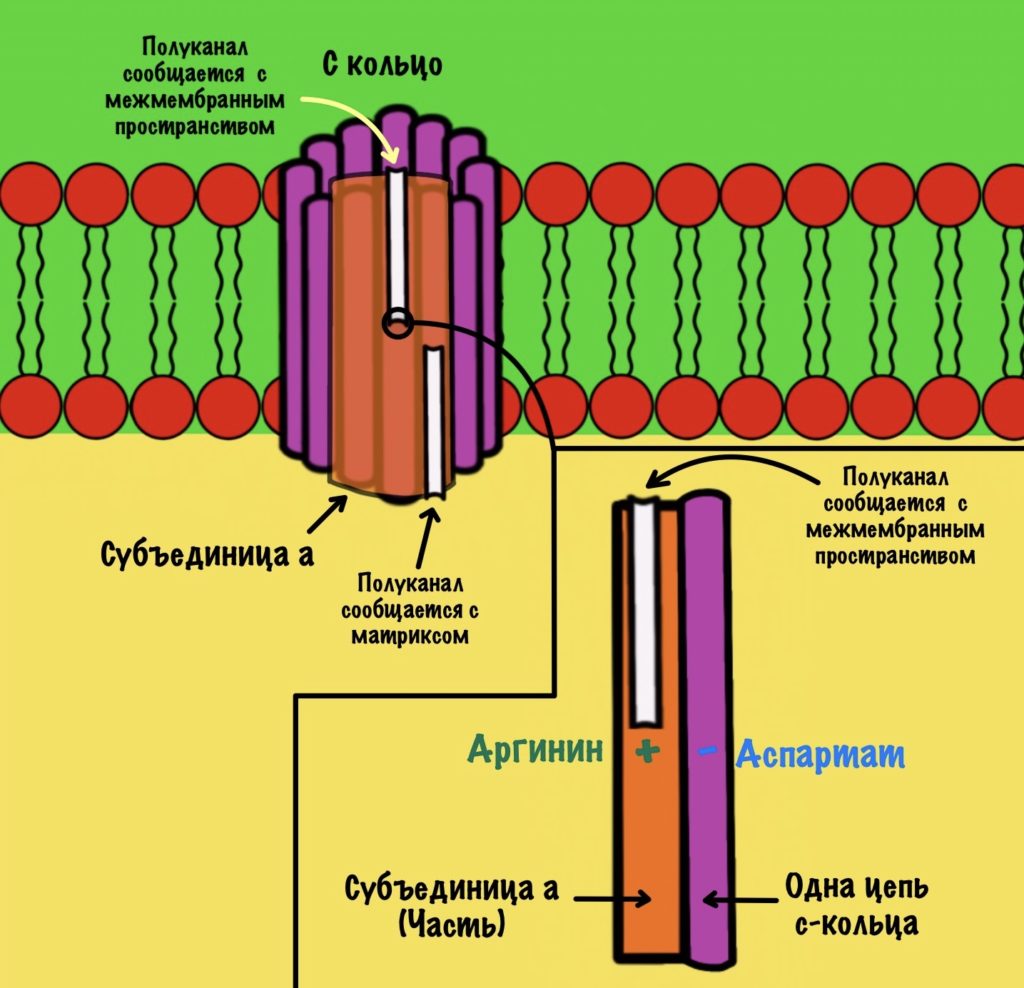
Теперь нужно понять механизм вращения ножки. Субъединица a связана с кольцом из 10-14 субъединиц c. Напоминаю, что в а-субъединице есть полуканалы для протонов.
Если посмотреть на этот полуканал, то мы увидим кое-что интересное. В конце полуканала находится аргинин от субъединицы a. Это положительно-заряженная аминокислота. Аргинин взаимодействует с аспартатом из субъединицы c. Аспартат — это отрицательно-заряженная аминокислота. Аспартат и аргинин взаимодействуют, как разноимённо-заряженные частицы. Немного разверну рисунок для удобства.
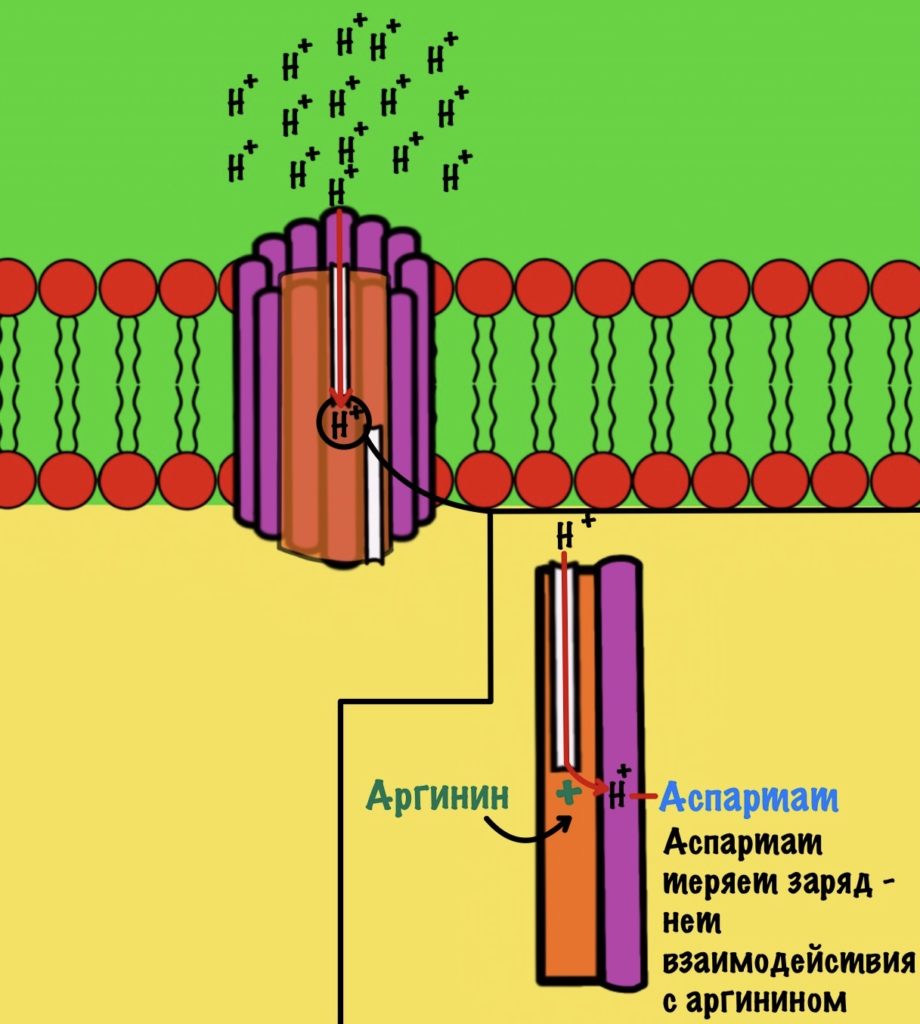
Протон пролетает через полуканал а субъединицы и присоединяется к аспартату. Из-за того, что у аспартата отрицательный заряд, а у протона положительный. Аспартат и аргинин перестают взаимодействовать друг с другом, так как аспартат потерял свой отрицательный заряд.
Это приводит к вращению всего кольца субъединиц c по часовой стрелке — на одну субъединицу c. При этом вращается и ножка гамма-эпсилон, что приводит к конформационным изменениям в гексамере. Ну это тот,который синтезирует АТФ.
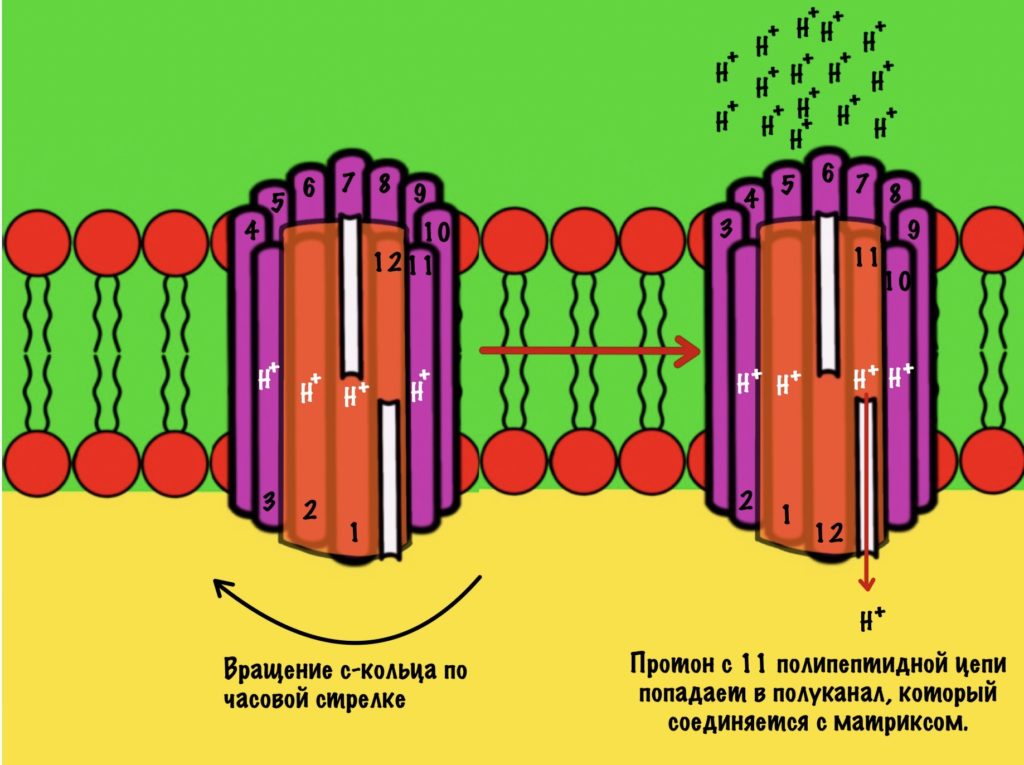
11 субъединица взаимодействует с каналом, который соединяется с матриксом. Аспартат этой субъединицы отдаёт протон водорода — он попадает в матрикс. Дальше 12 субъединица присоединит протон из межмембранного пространства, что приведёт к следующему вращению кольца. Теперь 11 субъединица присоединит протон и так далее… Удивительно, но природа придумала двигатель так давно!
Если мы хотим провести полное вращение c кольца — 360 градусов, то нам нужно присоединить к каждой субъединице с по одному протону. Так и происходит. Помните, что в кольце от 10 до 14 c субъединиц? Значит на полное вращение нам понадобится 10-14 протонов. В моём вот было 12 полипептидных цепей, значит для полного круга нужно присоединить 12 протонов.
При этом произойдёт полное вращение ножки — три раза по 120 градусов. Значит каждый гексамер изменит своё состояние три раза — три поворота по 120 градусов. Получается, что одно полное вращение с кольца, а с ним и ножки, приводит к синтезу трёх молекул АТФ.
Сколько протонов нужно для синтеза одной молекулы АТФ?
10-14 протонов на одно вращение на 360 градусов = 3 молекулы АТФ
Получается, что для синтеза одной молекулы АТФ нужно от 3,33 до 4,66 протонов. Или в среднем — 4 протона.
Для синтеза одной молекулы АТФ нужно 4 протона.
При транспорте электронов с НАДH было получено 10 протонов = 2,5 молекулы АТФ.
При транспорте электронов с ФАДH2 было получено 6 протонов = 1,5 молекулы АТФ.
Так что теперь вы не поведётесь на то, что пишут в некоторых учебниках: «Из одного НАДH синтезируется 3 молекулы АТФ, а из одного ФАДH2 — 2 молекулы АТФ. Поздравляю!
Нарушение работы окислительного фосфорилирования
Ух, ну и системку мы разобрали. Согласитесь, что она очень сложная. Но при этом отлично справляется со своей работой — в норме. Но давайте немного отойдём от нормы. Время поиграть в доктора!
Представьте себе, что у пациента анемия — снизились эритроциты и/или гемоглобин. Меньше кислорода поступает в ткани и органы, а значит в митохондриях дела плохи. Электроны не могут попасть на свой финальный акцептор — кислород. Электроны застревают в комплексах дыхательной цепи. Они не создают градиент концентрации, поэтому нет синтеза АТФ. Пациент с анемией чувствует общую слабость именно из-за этого.
В организме истощился запас железа. Тут два патологических механизма. Дефицит железа приводит к снижению гемоглобина, а дальше вы уже знаете. Железо идёт не только на синтез гемоглобина, но и цитохромов — компонентов третьего и четвёртого комплекса дыхательной цепи. Если нет акцепторов электронов, то нет и окислительного фосфорилирования.
Дефицит меди. Медь входит в четвёртый комплекс дыхательной цепи, поэтому если её не будет, то комплекс перестанет работать.
Дефицит белка в рационе или нарушение его всасывания. Причин множество, но нас они не интересуют. Белок используется для синтеза собственных белков организма — ферментов и комплексов дыхательной цепи.
Гиповитаминозы. Коснёмся только двух, хотя почти каждый гиповитаминоз будет приводить к нарушениям. НАД+ и ФАД — это производные витаминов. НАД+ — никотиновой кислоты (витамин B3). ФАД — рибофлавина (витамин B2). Если словить два этих гиповитаминоза, то у нас просто не будет начальных акцепторов электронов. Так что электрончики не смогут запрыгнуть на дыхательную цепь с НАДH и ФАДH2.
Дефицит углеводов и липидов в рационе/нарушение их всасывания. Тут всё понятно, если нет субстратов, то откуда взять электроны?
Разобщители окислительного фосфорилирования
Мы договорились, что работа дыхательной цепи сопряжена с синтезом АТФ. Но есть молекулы, которые разобщают эти два процесса. Как? Разобщители — это жирорастворимые молекулы, которые могут проходить через внутреннюю мембрану митохондрий. Но если бы они только проходили! Эти молекулы присоединяют к себе протоны, а потом переносят их в матрикс митохондрии. Они мешают митохондрии создать градиент концентрации протонов, который так важен для синтеза АТФ.
В итоге получается так, что дыхательная цепь работает, но синтеза АТФ нет. Энергия транспорта электронов рассеивается в виде тепла. Я хочу, чтобы вы помнили о разобщителях, которые встречаются в нашем организме: жирных кислотах, неконъюгированном билирубине и трийодтиронине. Это не значит, что в норме они сильно влияют на работу окислительного фосфорилирования. Но при патологических состояниях…
У пациента с ожирением в крови повышается количество жирных кислот. Чем больше жирных кислот, тем больше шанс, что они попадут в митохондрии и повлияют на окислительное фосфорилирование.
Некоъюгированный билирубин повышается при гемолизе эритроцитов или нарушениях функции печени. Его накопление приводит к разобщению работы дыхательной цепи и синтеза АТФ. Представьте какую слабость чувствует пациент при гемолизе эритроцитов — в митохондрии не попадает кислород, да ещё непрямой билирубин вызывает разобщение окислительного фосфорилирования. Мрак.
Трийодтиронин повышается при тиреотоксикозе. Поэтому эти пациенты чувствуют жар и быстро теряют в весе — энергия переноса электронов рассеивается в виде тепла.
Мы и сами можем вызывать разобщение, назначив дикумарин — антикоагулянт непрямого действия. Его побочный эффект — это разобщение окислительного фосфорилирования.
Подсчёт АТФ: сколько молекул можно получить с помощью окислительного фосфорилирования?
Теперь, когда мы понимаем все механизмы, можно подсчитать сколько АТФ было получено в ходе катаболизма одной молекулы глюкозы.
В ходе гликолиза было получено две молекулы НАДH. Они транспортируются в матрикс митохондрий — ведь гликолиз шёл в цитоплазме. Для этого используются челночные механизмы. Глицеролфосфатный челнок переносит электроны с НАДH на ФАД — образуется ФАДH2. Малат-аспартатный переносит электроны на митохондриальный НАД+ — переходит в НАДH. Теперь понимаете, почему это важно? Будет получено разное количество АТФ.
Остальные восстановленные коферменты уже находятся в митохондрии, поэтому их электроны можно сразу отправить на дыхательную цепь. Допустим, что мы использовали малат-аспартатный челнок, тогда в матриксе митохондрии находятся: 10 молекул НАДH и 2 молекулы ФАДH2.
10 х 2,5 АТФ + 2 х 1,5 АТФ = 28 молекул АТФ.
Если мы использовали глицеролфосфатный челнок, то в матриксе находятся: 8 молекул НАДH и 4 молекулы ФАДH2.
8 x 2,5 АТФ + 4 х 1,5 АТФ = 26 молекул АТФ.
В ходе окислительного фосфорилирования из одной молекулы глюкозы синтезируется 28 или 26 молекул АТФ.
Но это не общий выход катаболизма глюкозы, потому что в нём были и другие реакции синтеза АТФ — субстратного фосфорилирования. Если вспомнить гликолиз, то там были две таких реакции.
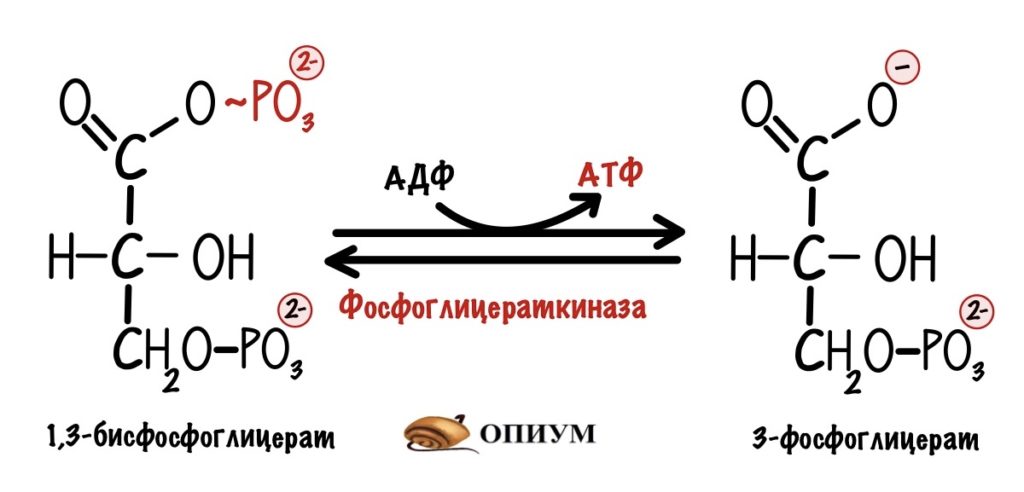
И вот вторая:
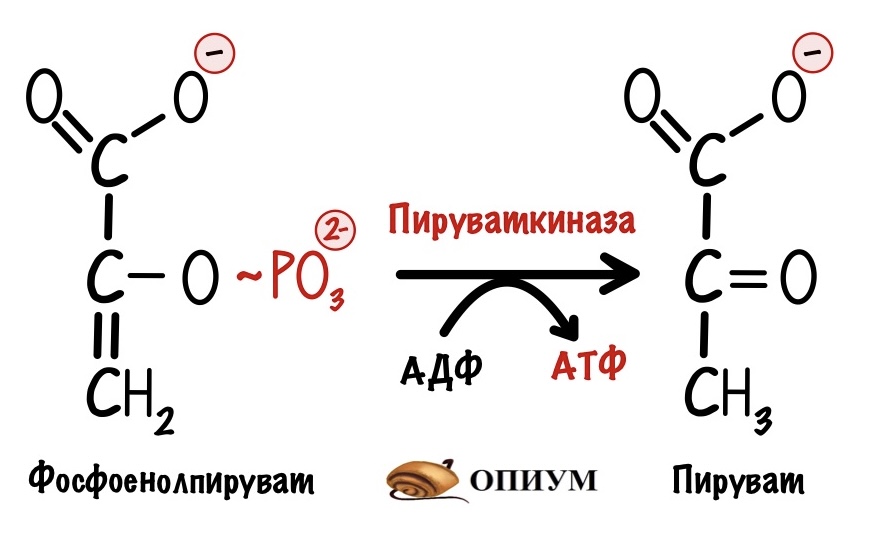
Мы видим, что из одной триозы образуется ещё 2 молекулы АТФ. Но в ходе гликолиза были образованы две триозы, поэтому всего там было получено 4 молекулы АТФ. Но это ещё не всё…
На активацию молекулы глюкозы мы затратили 2 молекулы АТФ — первая и третья реакции гликолиза.
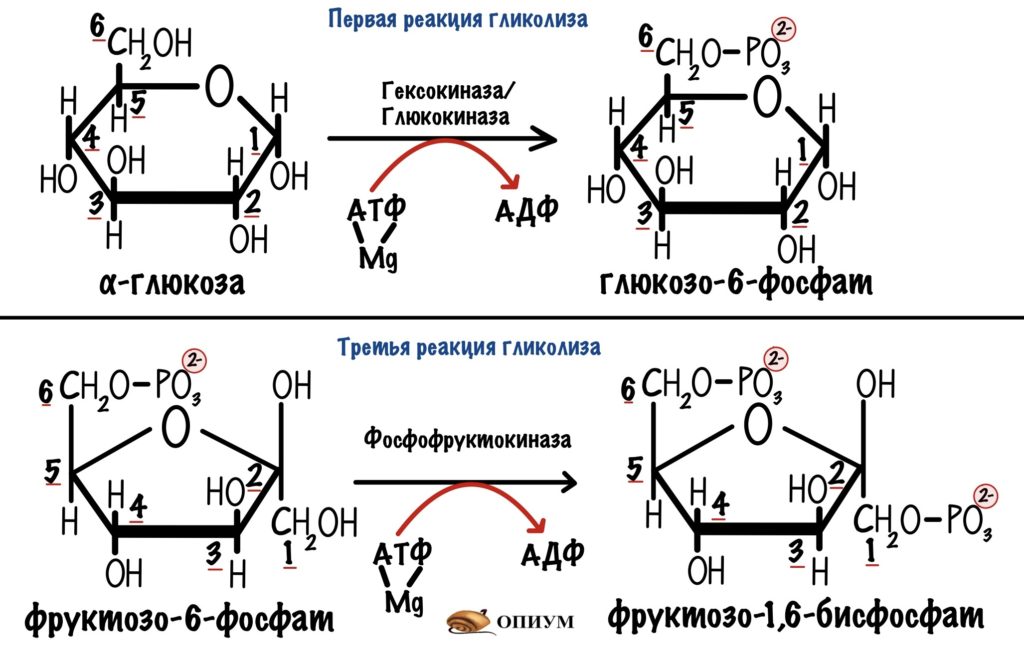
Так что общий выход гликолиза — 2 молекулы АТФ. Но и это ещё не всё!
Вспоминаем цикл трикарбоновых кислот, где были получены ещё две молекулы ГТФ. ГТФ равно АТФ, так как можно легко их переводить друг в друга.

Сделаем общий подсчёт. Гликолиз = 2 АТФ и 2 НАДH. Окислительное декарбоксилирование пирувата = 2 НАДH. ЦТК = 6 НАДH + 2 ФАДH2 + 2 ГТФ (АТФ). Итого:
2 АТФ + 10 НАДH + 2 ФАДH2 + 2 ГТФ (АТФ) = 2 + 25 + 3 + 2 = 32 молекулы АТФ
Это для малат-аспартатного челнока. Если будет использован глицеролфосфатный, то:
2 АТФ + 8 НАДH + 4 ФАДH2 + 2 ГТФ (АТФ) = 2 + 20 + 6 + 2 = 30 молекул АТФ
Из одной молекулы глюкозы образуется 32 молекулы АТФ, либо 30. Всё зависит от челнока, который был использован клеткой.
Вы здесь
Синонимы к словосочетанию «окислительное фосфорилирование»
Синонимы для словосочетания «окислительное фосфорилирование» не найдены.
-
Его называют окислительным фосфорилированием.
-
На кристах содержатся комплексы дыхательных ферментов, необходимые для окислительного фосфорилирования.
-
Известно, что в результате процессов гликолиза в печени, мышцах и некоторых других тканях в ходе так называемого цикла трикарбоновых кислот и окислительного фосфорилирования, на 1 моль глюкозы образуется 33 молекулы аденозин-трифосфорной кислоты (АТФ) — основного энергетического субстрата, используемого во всех жизненных «отправлениях» организма.
О новом в эволюции жизни на Земле, 2016
-
Являясь разобщителями окислительного фосфорилирования, ненасыщенные жирные кислоты ускоряют процессы окисления в митохондриях тканей и тем самым регулируют избыточное отложение триглицеринов (жиров).
-
Тканевое дыхание сопряжено с окислительным фосфорилированием внутри митохондриальной мембраны, благодаря которой возникает движущая сила — протонный градиент.